 2022, Vol. 49
2022, Vol. 49扩展功能
文章信息
- 邢浩, 杜 古尔·卫卫, 薛娜娜, 宋文娟, 赵莉, 戚冉
- XING Hao, Duguer Weiwei, XUE Na’na, SONG Wenjuan, ZHAO Li, QI Ran
- 哈密露天煤矿不同环境介质微生物群落特征分析
- Microbial community characteristics of different environmental media in Hami open-pit coal mine
- 微生物学通报, 2022, 49(11): 4525-4537
- Microbiology China, 2022, 49(11): 4525-4537
- DOI: 10.13344/j.microbiol.china.220283
-
文章历史
- 收稿日期: 2022-03-21
- 接受日期: 2022-05-11
- 网络首发日期: 2022-05-25

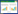




2. 新疆农业大学资源与环境学院, 新疆 乌鲁木齐 830052;
3. 中国科学院新疆生态与地理研究所, 新疆 乌鲁木齐 830011;
4. 中国地质调查局自然资源综合调查指挥中心, 北京 100055
2. College of Resources and Environment, Xinjiang Agricultural University, Urumqi 830052, Xinjiang, China;
3. Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Urumqi 830011, Xinjiang, China;
4. Command Center of Integrated Survey of Natural Resources, China Geological Survey, Beijing 100055, China
煤炭是全球第二大能源消耗来源。根据地形特征和煤层厚度等因素,煤矿开采包括地表(露天)开采和地下(井下)开采两种方式。煤炭生产过程中会产生大量的粉尘,采煤粉尘是一种复杂、非均匀、由不同粒径大小的颗粒物组成的混合物,其主要成分包括各种矿物、有机化合物及重金属等污染物[1]。采煤粉尘通过远距离传输和干湿沉降作用引发一系列诸如土壤重金属污染等生态环境问题[2-6]。采煤粉尘中的可吸入细颗粒物(PM2.5)引发的人体健康问题更是不容忽视[7]。
在矿区土壤环境中,微生物群落是土壤中元素循环及转化的重要参与者,在维持土壤生态平衡中扮演重要角色[8-9]。微生物群落结构与多样性可作为反映矿区土壤结构和质量的敏感性指标[10]。土壤重金属污染在改变土壤结构与功能的同时会引起土壤微生物群落结构的改变[11]。土壤中部分微生物可通过自身代谢活动对重金属具有吸附、吸收或转化功能,从而降低重金属的生态毒性[12]。然而,关于干旱区煤电能源基地土壤微生物群落特征研究还少有报道,仅有少数学者研究了露天煤矿围岩土壤微生物群落组成[13]、微生物群落稳定性对采矿活动的响应[14]。采煤粉尘及土壤风蚀产生的细颗粒物是煤矿区大气颗粒物的主要来源[7],微生物作为土壤环境的重要组成部分,其在PM2.5颗粒物中是否具有富集效应还不清楚,关于矿区PM2.5颗粒物的微生物群落组成还未见报道。因此,研究矿区大气PM2.5颗粒物的微生物组成对揭示PM2.5对人体健康的影响具有重要意义。
新疆维吾尔自治区哈密市南湖乡矿集区是我国重要的煤炭基地,煤矿开采引发了矿区土壤环境一定程度的重金属污染。微生物对重金属在土壤中的迁移和形态转化发挥重要作用[12],因此,开展矿区微生物生态研究对矿区污染生态影响的调查和恢复、矿区污染环境修复和生态保护具有非常重要的意义。本文以新疆维吾尔自治区哈密市南湖乡集露天开采和燃煤发电为一体的七号煤矿区为研究对象,基于Illumina NovaSeq测序平台,采用双末端测序的方法同步研究了新疆哈密露天煤矿区粉尘、土壤和PM2.5颗粒物中真菌和细菌的群落组成和功能特征,以期为干旱区露天煤矿土壤生态环境恢复、PM2.5颗粒物的健康风险评估及潜在功能微生物资源挖掘提供参考依据。
1 材料与方法 1.1 研究区概况研究区位于新疆维吾尔自治区哈密市南湖乡矿集区七号煤矿(93°19′06−93°29′00′′E,42°18′00′′−42°24′30′′N,海拔510−630 m),地处吐鲁番-哈密盆地东南部。区域水资源匮乏、气候干旱,地表类型属戈壁荒漠区,地表生物丰富度低,生态环境极为脆弱。区域内风沙、干热、低温寒潮等自然灾害多发。区内常年多风,风向以东北风为主,平均风速为3−4 m/s,最大风速可达23.5 m/s。七号煤矿是“西电东送”哈密南−郑州高压直流输电工程的配套煤源项目,是新疆煤炭工业重点开发项目。煤矿开采、存储等人为因素引发矿区内局部区域的土壤重金属污染和空气粉尘污染问题。
1.2 采样点布设与样品采集土壤样本采集自七号煤矿(中煤国投)电厂粉煤灰堆放区上风向(S1)及下风向(S2)土壤、露天采坑区上风向(S4)及下风向(S3)土壤、洗煤水排放场土壤(S7);粉尘样本采集自电厂粉煤灰堆场(S5)和露天采坑周边地表堆积的煤粉尘(S6);土壤样本采用五点混合法(100 m×100 m)采集地表表层(0−15 cm)土壤,土壤类型为棕漠土;粉尘样本采用五点混合法采集电厂粉煤灰和露天采坑煤粉尘的表层(0−15 cm)粉尘样本,取样后于−20 ℃保存待测。大气PM2.5颗粒物样本(P1−P4)采集点与土壤样本采集点(S1−S4)相对应(图 1)。使用便携式PM2.5空气采样器(Airmetrics公司)和石英滤膜(Whatman公司)采集PM2.5颗粒物样本,采样口高度距离地面1.8 m。采样器流量设置为5 L/min。24 h不间断采集,采集时长为3−7 d。采集完PM2.5颗粒物样本的滤膜于−20 ℃保存待测。

|
| 图 1 采样点分布图底图 Figure 1 The schematic diagram of sampling location. 为研究区2019年1: 50 000 Wordview-2遥感影像 The background picture is the 1: 50 000 Wordview-2 remote sensing image of study area in 2019. |
|
|
使用FastDNA® Spin Kit for Soil (MP公司)提取土壤和颗粒物样本的DNA,扩增质检合格的DNA样本。细菌多样性分析采用16S rRNA基因的V3−V4可变区序列引物341F (5′-CCTAC GGGNGGCWGCAG-3′)和805R (5′-GACTACHV GGGTATCTAATCC-3′)[15];真菌多样性分析采用ITS序列引物ITS1F (5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4R (5′-TCCTCCGCTTATTGATA TGC-3′)。PCR扩增产物经纯化、定量和均一化形成测序文库,建好的文库经质检合格后用Illumina MiSeq进行测序。
使用Trimmomatic V0.33软件,对测序得到的Raw Reads进行过滤;然后使用Cutadapt 1.9.1软件进行引物序列的识别与去除,得到不包含引物序列的Clean Reads;使用USEARCH V10软件,通过overlap对每个样品的Clean Reads进行拼接,然后根据不同区域的长度范围对拼接后数据进行长度过滤;使用UCHIME V4.2软件,鉴定并去除嵌合体序列,得到最终有效数据;按照97%相似性对非重复序列(不含单序列)进行聚类成为可操作分类单元(operational taxonomic units,OTU),由此得到OTU代表序列。采用RDP classifier贝叶斯算法对97%相似水平的OTU代表序列进行分类学分析(置信度阈值为80%)。研究所使用比对数据库为:细菌:Silva (http://www.arb-silva.de)[16];真菌:Unite (http://unite.ut.ee/index.php)[17]。
1.4 数据分析所有的数据分析均在R语言(V4.0.0)中完成。在未加特殊说明的情况下,文中所用图片均通过“ggplot2”包完成绘制[18]。其中,使用R包“picante”计算细菌群落的α多样性[19],使用“spaa”包计算不同样本微生物群落的生态位[20]。使用“vegan”包中的vegdist()函数计算得出样品间Bray-Curtis距离,使用hclust()函数进行非加权平均聚类分析(unweighted pair-group method with arithmetic means,UPGMA)[21]。
2 结果与分析 2.1 煤矿区真菌群落结构与组成从研究样本中得到79 741条优质真菌序列,1 971个OTU,分布在14个门中。在所有可注释的OTU中,63.53%−75.46%可归为子囊菌门(Ascomycota),11.29%−18.91%为担子菌门(Basidiomycota),4.03%−7.10%为未注释OTU (unclassified),其他11个门的平均相对丰度均低于5% (图 2)。在属水平上,优势真菌属有10个,包括曲霉属(Aspergillus)、镰刀菌属(Fusarium)、枝孢菌属(Cladosporium)、被孢霉属(Mortierella)、链格孢属(Alternaria)、青霉菌属(Penicillium)、毛葡孢属(Botryotrichum)、毛壳属(Chaetomidium)、红菇属(Russula)及拟青霉属(Paecilomyces)。图 2结果显示所有样本可分为两大支,其中S2、S3、S5、P1、P3、P4聚为一枝,S1、S4、S6、S7、P2聚为一支。

|
| 图 2 门水平下各样本中真菌群落相对丰度及其聚类图 Figure 2 Cluster analysis of fungal communities plot at the phylum level. |
|
|
从数量上来看,露天采坑区样本的真菌OTU总数显著高于电厂粉煤灰堆放区,而且同一工作区不同类型样本的真菌OTU总数存在明显差异。在露天采坑区,真菌OTU总数排序为土壤 > 大气PM2.5颗粒物 > 煤粉尘;在电厂粉煤灰堆放区,大气PM2.5颗粒物中的真菌OTU最高,其次是土壤,粉煤灰最低。从水平分布来看,大气PM2.5颗粒物、粉煤灰区(仅包括土壤和粉煤灰)与露天采坑区(仅包括土壤和煤粉尘)共有OTU总数为603个。然而,大气PM2.5颗粒物中可检出238个特有OTU,露天采坑区可检出339个,粉煤灰区可检出156个。
基于OTU水平的α多样性分析得出表征微生物群落丰度的Chao1指数和表征微生物群落多样性的Shannon指数。结果显示,露天采坑区真菌群落Chao1指数最小,为900.93±109.87;粉煤灰区Chao1指数为1 021.67±79.50,PM2.5颗粒物的Chao1指数最大,为1 088.39±99.51。但是,露天采坑区真菌群落的Shannon指数最大,为7.95±0.22;粉煤灰区Shannon指数为7.70±0.11,PM2.5颗粒物的Shannon指数最小,为7.69±0.17。统计分析表明,煤矿区土壤及PM2.5颗粒物在真菌群落丰度和多样性上均无显著性差异。对各样本真菌群落的生态位宽度计算得出,露天采坑区生态位宽度为2.98±0.09,粉煤灰区为3.02±0.07,PM2.5颗粒物为2.94±0.03,三者无显著性差异。
2.2 煤矿区真菌群落功能类群特征分析基于FUNGuild数据库,对注释好的真菌OTU进行潜在营养类型解析得出64类不同的功能分组。差异分析结果表明,所有功能类型在露天采坑区土壤、粉煤灰区土壤及PM2.5颗粒物之间不存在显著差异。两两组间差异分析得出,露天采坑区土壤和PM2.5颗粒物真菌群落功能无显著差异。露天采坑区和粉煤灰区土壤真菌群落之间发现3种不同功能类型具有显著性差异(P < 0.05) (图 3A),这3种类群均为腐生营养型,即可通过降解死亡的宿主细胞来获取营养。粉煤灰区土壤及PM2.5颗粒物之间也发现一类腐生营养型类群,其在粉煤灰区土壤中的丰度显著高于PM2.5颗粒物(P < 0.05) (图 3B)。

|
| 图 3 真菌功能类群差异性分析 Figure 3 Differential analysis of fungal functional groups. Welchʼs t-test, P < 0.05. |
|
|
从研究样本中可得到77 095条优质细菌序列,2 143个OTU,分布在32个门中。在所有可注释的OTU中,变形菌门(Proteobacteria)的相对丰度最高,为20.81%−33.80%;其次是放线菌门(Actinobacteria),相对丰度为5.42%−38.44%;厚壁菌门(Firmicutes),相对丰度为10.82%;酸杆菌门(Acidobacteria),相对丰度为10.78%;拟杆菌门(Bacteroidetes),相对丰度为6.36%;绿弯菌门(Chloroflexi),相对丰度为5.38%,其余26门的平均相对丰度均低于5% (图 4)。UPGMA树状图(图 4)结果显示细菌样本的聚类结果与真菌显著不同,S4和S6聚为一支,其余样本聚为一大支,其中P1、P2、P4及S3可单独聚为一小支。

|
| 图 4 门水平下各样本细菌群落相对丰度及聚类图 Figure 4 Relative abundance and cluster analysis of bacterial communities plot at the phylum level. |
|
|
从数量上来看,露天采坑区样本的细菌OTU总数略高于电厂粉煤灰堆放区,同一工作区不同类型样本的细菌OTU总数存在明显差异。在露天采坑区,细菌OTU总数排序为大气PM2.5颗粒物 > 土壤 > 煤粉尘;在电厂粉煤灰堆放区,土壤中的真菌OTU最高,其次是大气PM2.5颗粒物,粉煤灰最低。从水平分布来看,大气PM2.5颗粒物、粉煤灰区(仅包括土壤和粉煤灰)与露天采坑区(仅包括土壤和煤粉尘)共有OTU总数为1 767个。然而,大气PM2.5颗粒物中可检出10个特有细菌OTU,露天采坑区可检出83个,粉煤灰区可检出15个独有细菌OTU。
基于OTU水平的α多样性分析结果显示,PM2.5颗粒物的细菌丰富度指数Chao1值最高,为1 627.92±106.60,其次是露天采坑区和粉煤灰区,Chao1值分别为1 446.66±282.19和1 493.70±391.87。露天采坑区土壤细菌群落的Shannon指数为7.18±0.94,粉煤灰区为8.05±0.82,PM2.5颗粒物为8.89±0.41。综上所述,矿区PM2.5颗粒物的细菌丰富度和群落多样性均高于土壤和粉尘,但不同工作区的土壤细菌群落多样性无显著性差异。各样本的细菌群落生态位宽度计算结果发现,露天采坑区生态位宽度为4.01±0.19,粉煤灰区为4.12±0.27,PM2.5颗粒物的细菌群落生态位宽度为4.45±0.06。方差分析结果表明,PM2.5颗粒物的细菌群落生态位宽度最大,显著高于露天采坑区,但其与粉煤灰区无显著差异。
通过线性判别分析(linear discriminant analysis,LDA)可得到样本中不同分类水平中存在组间差异的细菌进化分枝树图。如图 5所示,矿区内不同工作区土壤及PM2.5颗粒物的细菌群落中的特征物种的统计结果表明,母链菌纲(Blastocatellia_subgroup_4)、伯克氏菌科(Burkholderiaceae)及δ变形菌目(Deltaproteobacteria)细菌为露天采坑区土壤中的特征物种。绿弯菌门(Chloroflexi)类细菌则为粉煤灰区土壤细菌群落的特征物种。此外,PM2.5颗粒物细菌群落中的特征物种为酸杆菌门(Acidobacteria)、硝化螺旋菌门(Nitrospirae)及变形菌门(Proteobacteria)。

|
| 图 5 存在组间差异的细菌进化分支树图 Figure 5 The taxonomic branching tree of bacterial evolution with intergroup difference. |
|
|
将矿区细菌群落与FAPROTAX数据库进行比对,可鉴定出56个生态功能簇。其中包括碳(C)循环相关功能类群(如甲烷营养型)、氮(N)循环相关功能类群(如硝化反硝化作用)、硫(S)循环相关功能类群(如S的氧化还原)及部分其他元素循环相关的功能类群(如铁的氧化和还原)。总体来看,甲烷营养型细菌类群在露天采坑区的丰度显著高于粉煤灰区和PM2.5颗粒物(P < 0.05)。但是,粉煤灰区土壤中几丁质酶类细菌类群的丰度显著高于露天采坑区和PM2.5颗粒物(P < 0.05)。两两组间差异性分析结果表明,在露天采坑区和粉煤灰区土壤中,具尿素分解功能的细菌类群丰度存在显著差异(P=0.029),露天采坑区高于粉煤灰区(图 6A)。好氧氨氧化细菌和纤维素水解功能菌的丰度在露天采坑区和PM2.5颗粒物中存在显著差异(P < 0.05),露天采坑区均低于PM2.5颗粒物(图 6B)。此外,部分具有寄生功能的类群、甲醇氧化以及甲基营养型细菌类群在PM2.5颗粒物中的丰度要显著高于粉煤灰区土壤(P < 0.05) (图 6C)。

|
| 图 6 细菌功能类群差异分析 Figure 6 Differential analysis of bacterial functional groups. Welchʼs t-test, P < 0.05. |
|
|
目前国内外关于矿区土壤微生物多样性的研究比较多,如我国学者对铜矿区[22-23]、铅锌银尾矿区[8]土壤微生物群落多样性开展了研究,均证实重金属元素会降低土壤微生物多样性,改变微生物群落结构。露天煤矿开采区是一种典型的人为扰动形成的极端环境,其特征是无机(如重金属)和有机(如多环芳烃)污染物易导致土壤环境地球化学特征不稳定,引起微生物的氧化应激反应[24]。Illumina MiSeq测序技术被广泛应用于环境微生物多样性研究,然而16S rRNA基因的组内异质性可能导致原核生物多样性被高估[25]。因此,测序引物的选择对于分析微生物的多样性至关重要,有研究得出16S rRNA基因V3−V4和V4−V5可变区引物组在代表总微生物群落组成从门到属分类水平方面具有高度相似性[26]。本研究分别采用16S rRNA基因V3−V4可变区序列和引物ITS,研究了哈密七号露天煤矿不同环境介质(包括土壤、粉尘和PM2.5颗粒物)的细菌和真菌群落组成及多样性。研究发现,矿区优势真菌群落为子囊菌门(Ascomycota),优势细菌群落为变形菌门(Proteobacteria)和放线菌门(Actinobacteria)。该结果与内蒙古露天煤矿土壤微生物群落的结构组成[14]一致。子囊菌门和变形菌门分别是自然环境中分布最为广泛、丰度最高的真菌及细菌类群[27-28]。露天煤矿采坑和矿井周围的土壤中会产生大量的煤基质,其主要由芳香烃、脂肪族和木质素衍生的大分子复杂混合物组成[29]。有研究发现,上述优势菌群能够参与产甲烷菌的代谢,以促进生物甲烷的产生和煤的降解[30]。例如,子囊菌门中的镰刀菌属(Fusarium sp.)对多环芳烃(polycyclic aromatic hydrocarbons,PAH)具有高效降解作用[31],放线菌门中的木质素分解链霉菌属(ligninolytic Streptomyces)可通过分泌独特的碱性产物来生物降解褐煤和烟煤[32]。子囊菌门的茎点霉属细菌可通过胞外沉淀作用[33]、球囊霉可通过原生质膜的离子交换作用来提高对重金属的耐受性[34]。部分变形菌门的细菌可通过编码金属硫蛋白进而结合重金属,或通过胞外聚合物吸收、转化重金属,从而降低重金属的毒性[35]。因此,在重金属等污染物产生的氧化应激条件下,子囊菌门(Ascomycota)、变形菌门(Proteobacteria)和放线菌门(Actinobacteria)比其他微生物具备更好的生存能力,它们在矿区污染生态修复中发挥重要作用[36]。
人类活动,如露天煤矿开采扰动改变了土壤的物理和化学性质,从而改变了土壤微生物群落的结构。自然条件下煤覆盖层的属性(如硫含量)[37],土壤环境pH[38]对矿区土壤细菌群落组成和多样性具有重要影响。有研究发现,矿区高硫煤(干燥基全硫Std含量 > 3.00%的煤)覆盖能够显著改变细菌群落组成,优势菌为变形菌门嗜酸菌属(Acidiphilium)和厚壁菌门硫化杆菌属(Sulfobacillus)[37]。本研究中,嗜酸菌属(Acidiphilium)和硫化杆菌属(Sulfobacillus)的相对丰度较低,可能与该矿区煤层的低硫含量(平均Std为0.42%)属性和区域土壤呈碱性(pH均值为8.4)相关。部分学者研究发现,在重金属污染严重的有色金属矿区,如铜陵狮子山矿区[39]和汞污染严重的北京怀柔金矿区[40],土壤优势细菌群落也是变形菌门(Proteobacteria)和放线菌门(Actinobacteria),它们也是矿区土壤重金属抗性细菌的主要来源。上述结果表明,变形菌门(Proteobacteria)和放线菌门(Actinobacteria)对矿区环境具有广泛的环境适应性。
综上所述,在以往的研究中,土壤是矿区微生物研究的主要对象。本研究发现矿区微生物群落组成及多样性与环境样品类型密切相关。从区域整体来看,土壤、粉尘和PM2.5颗粒物的群落组成无显著差异,这可能是由于粉尘多样化的扩散方式导致的矿区内微生物群落结构的趋同效应。生态位宽度差异分析发现,矿区PM2.5颗粒物的细菌生态位宽度较大,可能原因有两方面,一方面煤矿区粉尘中的细颗粒物在风力作用下实现远距离扩散,由点源污染转变为面源污染;另一方面,携带重金属等污染物的粉尘可通过大气干湿沉降进入地表环境,对区域生态系统造成破坏[5]。从环境样品类型来看,PM2.5颗粒物中细菌的丰富度和群落多样性均高于土壤和粉尘,但是真菌组成在各个环境介质之间无明显差异性。矿区大气PM2.5颗粒物与工人和周边居民的人体健康密切相关。据研究报道,煤矿区PM2.5颗粒物能够导致人类细胞染色体的丢失和断裂[41]。与土壤相比,PM2.5颗粒物中丰度较高的酸杆菌门(Acidobacteria)、硝化螺旋菌门(Nitrospirae)及变形菌门(Proteobacteria)也是环境中抗生素耐药基因的主要宿主,即潜在的耐药菌[42]。此外,本研究发现具有寄生功能的细菌类群在PM2.5颗粒物中存在富集。因此,矿区PM2.5颗粒中的细菌对人类健康可能具有更严重的影响。
基于微生物群落组成的功能类群预测分析可为理解不同环境及生态修复过程中微生物群落差异提供重要信息。有学者在矿区土壤生态恢复的过程中发现,添加有机肥和微生物菌剂的处理组中与碳、氮、磷、硫循环相关的功能微生物类群显著提升[43]。本研究发现,露天采坑区土壤、粉煤灰土壤及PM2.5颗粒物中均存在不同分类水平特征的细菌和真菌类群。其中,矿区土壤中腐生营养型类群显著高于PM2.5颗粒物;与碳循环相关的甲烷营养型细菌类群显著富集于露天采坑区,具几丁质分解功能的细菌类群富集于粉煤灰区土壤。这些功能微生物类群与其所在区域的生态环境密切相关,基于上述信息可以针对性筛选矿区不同生态功能区特异的微生物类群用于受损生态环境的修复。然而,关于这些特征类群微生物对生态环境质量会产生什么作用,尤其是PM2.5颗粒物中的细菌类群对人体健康的影响,还需进一步研究。此外,微生物群落与环境因子之间的微妙关系能够影响微生物群落的稳定性和生态系统水平过程。因此,为综合预测微生物对人为扰动的响应过程,未来还需深入研究影响微生物群落稳定性的环境驱动因素。
4 结论(1) 哈密露天煤矿七号矿区土壤和粉尘颗粒物中的优势真菌群落来源于子囊菌门(Ascomycota)和担子菌门(Basidiomycota)。大气PM2.5颗粒物、粉煤灰区(仅包括土壤和粉煤灰)与露天采坑区(仅包括土壤和煤粉尘)共有OTU总数为603个,特有OTU数分别为238、156和339个。
(2) 不同功能区(包括露天采坑和粉煤灰堆放区)的土壤与大气PM2.5颗粒物在真菌群落丰度、群落多样性和群落生态位宽度无显著性差异。矿区内的真菌特征功能类群为腐生营养型类群。
(3) 矿区的优势细菌群落来源于变形菌门(Proteobacteria)和放线菌门(Actinobacteria)。大气PM2.5颗粒物、粉煤灰区与露天采坑区共有OTU总数为1 767个,特有OTU数分别为10、15和83个。
(4) 大气PM2.5颗粒物的细菌丰富度和群落多样性均高于土壤和粉尘,但不同工作区的土壤细菌群落多样性无显著性差异。PM2.5颗粒物细菌群落生态位宽度显著大于露天采坑区和粉煤灰区。矿区内的细菌特征功能类群包括甲烷营养型类群、几丁质酶类细菌类群、好氧氨氧化细菌类群、纤维素水解功能菌群、甲醇氧化及甲基营养型细菌类群。
致谢: 本研究实验样本的获取得到中国科学院新疆生态与地理研究所环境污染与生态修复实验室的支持和帮助,在此表示衷心的感谢。| [1] |
Liu GJ, Vassilev SV, Gao LF, Zheng LG, Peng ZC. Mineral and chemical composition and some trace element contents in coals and coal ashes from Huaibei coal field, China[J]. Energy Conversion and Management, 2005, 46(13/14): 2001-2009. |
| [2] |
Hu XH, Zhang L, Kong LF, Wang TJ. Studies on pollution risks of heavy metals in dustfall, soil and plant around the opencast coal mines in eastern Junggar Basin in Xinjiang[J]. Environmental Protection of Xinjiang, 2020, 42(4): 24-32. (in Chinese) 胡潇涵, 张琳, 孔利锋, 王天娇. 新疆准东露天煤矿降尘、土壤以及植物中的重金属污染风险研究[J]. 新疆环境保护, 2020, 42(4): 24-32. DOI:10.3969/j.issn.1008-2301.2020.04.007 |
| [3] |
Li KJ, Gu YS, Li MZ, Zhao L, Ding JJ, Lun ZJ, Tian W. Spatial analysis, source identification and risk assessment of heavy metals in a coal mining area in Henan, Central China[J]. International Biodeterioration & Biodegradation, 2018, 128: 148-154. |
| [4] |
Fan Q, Li SY, Guan TL, Wu XX, Wang R, Ren LJ. The ecological effect on plant and soil around opencast coal mine from the mineral dust[J]. Northern Environment, 2013, 25(9): 104-108. (in Chinese) 樊强, 李素英, 关塔拉, 武晓霞, 王冉, 任丽娟. 露天煤矿生产中产生的粉尘对周边植物和土壤的影响[J]. 北方环境, 2013, 25(9): 104-108. |
| [5] |
Cheng HX, Li M, Zhao CD, Li K, Peng M, Qin AH, Cheng XM. Overview of trace metals in the urban soil of 31 metropolises in China[J]. Journal of Geochemical Exploration, 2014, 139: 31-52. DOI:10.1016/j.gexplo.2013.08.012 |
| [6] |
Li CC, Zhang GS, Yao F, Wang Y, Li HD. Assessment of soil heavy metal pollution in area of Xingjiang Zhundong wucaiwan surface coal mine[J]. Environmental Engineering, 2014, 32(7): 142-146. (in Chinese) 李长春, 张光胜, 姚峰, 王宇, 李昊东. 新疆准东煤田五彩湾露天矿区土壤重金属污染评估与分析[J]. 环境工程, 2014, 32(7): 142-146. DOI:10.13205/j.hjgc.201407031 |
| [7] |
Moreno T, Trechera P, Querol X, Lah R, Johnson D, Wrana A, Williamson B. Trace element fractionation between PM10 and PM2.5 in coal mine dust: implications for occupational respiratory health[J]. International Journal of Coal Geology, 2019, 203: 52-59. DOI:10.1016/j.coal.2019.01.006 |
| [8] |
Teng Y, Huang CY, Luo YM, Long J, Yao HY. Microbial activities and functional diversity of community in soils polluted with Pb-Zn-Ag mine tailings[J]. Acta Pedologica Sinica, 2004, 41(1): 113-119. (in Chinese) 滕应, 黄昌勇, 骆永明, 龙健, 姚槐应. 铅锌银尾矿区土壤微生物活性及其群落功能多样性研究[J]. 土壤学报, 2004, 41(1): 113-119. DOI:10.3321/j.issn:0564-3929.2004.01.019 |
| [9] |
Fan JX, Gao CH, Lu CD, Zhang Q, Jin DS, Li JH. Review on mine soil microorganism diversity research[J]. Journal of Shanxi Agricultural Sciences, 2010, 38(3): 55-58. (in Chinese) 范继香, 郜春花, 卢朝东, 张强, 靳东升, 李建华. 矿区土壤微生物多样性研究概述[J]. 山西农业科学, 2010, 38(3): 55-58. DOI:10.3969/j.issn.1002-2481.2010.03.17 |
| [10] |
Yi JX, Li J. Research progress on microbial diversity of reclaimed soil in mining area[J]. Agricultural Science-Technology and Information, 2018(11): 42-45. (in Chinese) 弋嘉喜, 李娟. 矿区复垦土壤微生物多样性研究进展[J]. 农业科技与信息, 2018(11): 42-45. DOI:10.15979/j.cnki.cn62-1057/s.2018.11.017 |
| [11] |
Yu FM, Yao YW, Xie DY, Wang XR, Lin JM, Liu Y, Liu KH, Li Y. Study on the soil microbial community structure associated with six land use in Siding mining area[J]. China Environmental Science, 2020, 40(5): 2262-2269. (in Chinese) 于方明, 姚亚威, 谢冬煜, 王雪茹, 林嘉敏, 刘媛, 刘可慧, 李艺. 泗顶矿区6种土地利用类型土壤微生物群落结构特征[J]. 中国环境科学, 2020, 40(5): 2262-2269. DOI:10.3969/j.issn.1000-6923.2020.05.049 |
| [12] |
Li MJ. Microbial diversity in mine environment with metal pollution[D]. Xi'an: Master's Thesis of Xi'an University of Architecture and Technology, 2016 (in Chinese) 李梦杰. 重金属污染矿区微生物多样性分析[D]. 西安: 西安建筑科技大学硕士学位论文, 2016 |
| [13] |
Jiang BH, Zhang B, Li L, Zhao Y, Shi Y, Jiang Q, Jia LP. Analysis of microbial community structure and diversity in surrounding rock soil of different waste dump sites in Fushun western opencast mine[J]. Chemosphere, 2021, 269: 128777. DOI:10.1016/j.chemosphere.2020.128777 |
| [14] |
Chen J, Nan J, Xu DL, Mo L, Zheng YX, Chao LM, Qu HT, Guo YQ, Li FS, Bao YY. Response differences between soil fungal and bacterial communities under opencast coal mining disturbance conditions[J]. CATENA, 2020, 194: 104779. DOI:10.1016/j.catena.2020.104779 |
| [15] |
Herlemann DP, Labrenz M, Jürgens K, Bertilsson S, Waniek JJ, Andersson AF. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea[J]. The ISME Journal, 2011, 5(10): 1571-1579. DOI:10.1038/ismej.2011.41 |
| [16] |
Quast C, Pruesse E, Yilmaz P, Gerken J, Glckner FO. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools[J]. Nucleic Acids Research, 2012, 41(D1): D590-D596. DOI:10.1093/nar/gks1219 |
| [17] |
Kõljalg U, Nilsson RH, Abarenkov K, Tedersoo L, Taylor AFS, Bahram M, Bates ST, Bruns TD, Bengtsson-Palme J, Callaghan TM, et al. Towards a unified paradigm for sequence-based identification of fungi[J]. Molecular Ecology, 2013, 22(21): 5271-5277. DOI:10.1111/mec.12481 |
| [18] |
Wickham H. ggplot2: Elegant Graphics for Data Analysis[M]. Springer, 2009
|
| [19] |
Kembel SW, Cowan PD, Helmus MR, Cornwell WK, Morlon H, Ackerly DD, Blomberg SP, Webb CO. Picante: R tools for integrating phylogenies and ecology[J]. Bioinformatics, 2010, 26(11): 1463-1464. DOI:10.1093/bioinformatics/btq166 |
| [20] |
Zhang JL, Ding Q, Huang JH. spaa: species association analysis[CP]. R package version 0.2, 2013, 1
|
| [21] |
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O'hara RB, Simpson GL, Solymos P, et al. Vegan: Community Ecology Package. R package version 2.2-0, 2014
|
| [22] |
Chen JW, Zhang H, Li JJ, Liu Y. Soil fungal community structure and functional diversity in a copper tailing dam and its surrounding areas[J]. Environmental Science, 2021, 42(4): 2056-2065. (in Chinese) 陈建文, 张红, 李君剑, 刘勇. 铜尾矿坝及其周边土壤真菌群落结构与功能多样性[J]. 环境科学, 2021, 42(4): 2056-2065. DOI:10.13227/j.hjkx.202008136 |
| [23] |
Li C, Jing JH, Liu JX, Chai BF. Driving factors of the dynamics of microbial community in a dam of copper mine tailings[J]. Environmental Science, 2018, 39(4): 1804-1812. (in Chinese) 李毳, 景炬辉, 刘晋仙, 柴宝峰. 铜尾矿库坝面土壤微生物群落动态的驱动因子[J]. 环境科学, 2018, 39(4): 1804-1812. |
| [24] |
Li XF, Huang LB, Bond PL, Lu Y, Vink S. Bacterial diversity in response to direct revegetation in the Pb-Zn-Cu tailings under subtropical and semi-arid conditions[J]. Ecological Engineering, 2014, 68: 233-240. DOI:10.1016/j.ecoleng.2014.03.044 |
| [25] |
Sun DL, Jiang X, Wu QL, Zhou NY. Intragenomic heterogeneity of 16S rRNA genes causes overestimation of prokaryotic diversity[J]. Applied and Environmental Microbiology, 2013, 79(19): 5962-5969. DOI:10.1128/AEM.01282-13 |
| [26] |
Fadeev E, Cardozo-Mino MG, Rapp JZ, Bienhold C, Salter I, Salman-Carvalho V, Molari M, Tegetmeyer HE, Buttigieg PL, Boetius A. Comparison of two 16S rRNA primers (V3−V4 and V4−V5) for studies of Arctic microbial communities[J]. Frontiers in Microbiology, 2021, 12: 637526. DOI:10.3389/fmicb.2021.637526 |
| [27] |
Spain AM, Krumholz LR, Elshahed MS. Abundance, composition, diversity and novelty of soil Proteobacteria[J]. The ISME Journal, 2009, 3(8): 992-1000. DOI:10.1038/ismej.2009.43 |
| [28] |
Al-Sadi AM. High fungal diversity and dominance by Ascomycota in dam reservoir soils of arid climates[J]. International Journal of Agriculture and Biology, 2017, 19(4): 682-688. DOI:10.17957/IJAB/15.0328 |
| [29] |
Strąpoć D, Mastalerz M, Dawson K, Macalady J, Callaghan AV, Wawrik B, Turich C, Ashby M. Biogeochemistry of microbial coal-bed methane[J]. Annual Review of Earth and Planetary Sciences, 2011, 39: 617-656. DOI:10.1146/annurev-earth-040610-133343 |
| [30] |
Wang BB, Wang YF, Cui XY, Zhang YM, Yu ZS. Bioconversion of coal to methane by microbial communities from soil and from an opencast mine in the Xilingol grassland of northeast China[J]. Biotechnology for Biofuels, 2019, 12: 236. DOI:10.1186/s13068-019-1572-y |
| [31] |
Zhao OY, Feng SD, Jia HB, Zhang XN, Wei S, Wei W, Yang ZX, Li YL. Biodegradation of high molecular weight polycyclic aromatic hydrocarbons mixture by a newly isolated Fusarium sp. and co-metabolic degradation with starch[J]. Polycyclic Aromatic Compounds, 2018, 38(1): 32-41. DOI:10.1080/10406638.2016.1143847 |
| [32] |
Gupta RK, Spiker JK, Crawford DL. Biotransformation of coal by ligninolytic Streptomyces[J]. Canadian Journal of Microbiology, 1988, 34(5): 667-674. DOI:10.1139/m88-110 |
| [33] |
Yuan HL, Li ZJ, Ying JY, Wang ET. Cadmium(II) removal by a hyperaccumulator fungus Phoma sp. F2 isolated from blende soil[J]. Current Microbiology, 2007, 55(3): 223-227. DOI:10.1007/s00284-007-0088-z |
| [34] |
González-Guerrero M, Melville LH, Ferrol N, Lott JNA, Azcón-Aguilar C, Peterson RL. Ultrastructural localization of heavy metals in the extraradical mycelium and spores of the arbuscular mycorrhizal fungus Glomus intraradices[J]. Canadian Journal of Microbiology, 2008, 54(2): 103-110. |
| [35] |
Chihomvu P, Stegmann P, Pillay M. Identification and characterization of heavy metal resistant bacteria from the Klip River[R]. Proceedings of the International Conference on Ecological, Environmental and Biological Sciences, WASET, Cape Town, South African. 2014, 2526
|
| [36] |
Sánchez-Castro I, Gianinazzi-Pearson V, Cleyet-Marel JC, Baudoin E, Van Tuinen D. Glomeromycota communities survive extreme levels of metal toxicity in an orphan mining site[J]. Science of the Total Environment, 2017, 598: 121-128. |
| [37] |
Dong SB, Li N, Xu JF, Li JQ. Influence of high-sulfur coal cover on composition and diversity of soil bacterial community in mining areas[J]. Microbiology China, 2021, 48(3): 710-721. (in Chinese) 董书博, 李宁, 许继飞, 李静泉. 矿区高硫煤覆盖对土壤细菌群落组成和多样性的影响[J]. 微生物学通报, 2021, 48(3): 710-721. |
| [38] |
Lauber CL, Hamady M, Knight R, Fierer N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale[J]. Applied and Environmental Microbiology, 2009, 75(15): 5111-5120. |
| [39] |
Zhao XQ, Sun Y, Huang J, Wang H, Tang D. Effects of soil heavy metal pollution on microbial activities and community diversity in different land use types in mining areas[J]. Environmental Science and Pollution Research International, 2020, 27(16): 20215-20226. |
| [40] |
Ji HB, Zhang Y, Bararunyeretse P, Li HX. Characterization of microbial communities of soils from gold mine tailings and identification of mercury-resistant strain[J]. Ecotoxicology and Environmental Safety, 2018, 165: 182-193. |
| [41] |
Espitia-Pérez L, Da Silva J, Espitia-Pérez P, Brango H, Salcedo-Arteaga S, Hoyos-Giraldo LS, De Souza CT, Dias JF, Agudelo-Castañeda D, Valdés Toscano A, et al. Cytogenetic instability in populations with residential proximity to open-pit coal mine in northern Colombia in relation to PM10 and PM2.5 levels[J]. Ecotoxicology and Environmental Safety, 2018, 148: 453-466. |
| [42] |
Xu LK, Campos LC, Canales M, Ciric L. Drinking water biofiltration: behaviour of antibiotic resistance genes and the association with bacterial community[J]. Water Research, 2020, 182: 115954. |
| [43] |
Yan X, Wang JL, Hu XW, Yu BB, Gao BW, Li ZL, Chen JH, Zhang MJ, Liu XY. Contrasting effects of microbial fertiliser and organic fertiliser on soil bacterial community in coal mine dump of Inner Mongolia[J]. Chemistry and Ecology, 2021, 37(4): 384-398. |