 2019, Vol. 46
2019, Vol. 46扩展功能
文章信息
- 张译兮, 张亚妮, 马艳玲, 罗威, 肖雅, 孔伟娜
- ZHANG Yi-Xi, ZHANG Ya-Ni, MA Yan-Ling, LUO Wei, XIAO Ya, KONG Wei-Na
- 铜绿假单胞菌中Ⅲ型分泌系统调控机制及针对性治疗的研究进展
- Progress in the regulation mechanism and targeted treatment of type Ⅲ secretion system in Pseudomonas aeruginosa
- 微生物学通报, 2019, 46(2): 362-373
- Microbiology China, 2019, 46(2): 362-373
- DOI: 10.13344/j.microbiol.china.180465
-
文章历史
- 收稿日期: 2018-06-08
- 接受日期: 2018-08-06
- 网络首发日期: 2018-08-10

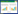




铜绿假单胞菌(Pseudomonas aeruginosa)又称绿脓杆菌,是一种适应性很强的革兰氏阴性条件致病菌。它在自然界中分布广泛,在人群中可以引起急性和慢性感染。铜绿假单胞菌易感人群主要有烧伤、呼吸道及肺部感染、癌症和艾滋病等免疫力低下患者,尤其在囊性纤维化(Cystic fibrosis,CF)患者中,该菌感染是最主要的致死原因[1-2]。Ⅲ型分泌系统(Type Ⅲ secretion system,T3SS)是铜绿假单胞菌急性感染过程中的关键毒性因子,通过结构复杂的针状分泌装置可将几种效应蛋白直接注入真核宿主细胞内,引起宿主细胞产生相应的病变,并帮助铜绿假单胞菌逃逸巨噬细胞的吞噬[3]。鉴于T3SS对铜绿假单胞菌致病过程的重要性,目前越来越多的研究者对这一系统的结构、功能及调控机制进行了研究,同时针对该系统的小分子抑制剂及疫苗的研发也为控制和预防铜绿假单胞菌的感染提供了新的治疗手段。本文结合近年来铜绿假单胞菌的最新研究结果以及我们课题组在T3SS方面的研究工作,阐述了T3SS的具体调控机制和针对性治疗策略。
1 铜绿假单胞菌T3SS的结构及功能铜绿假单胞菌T3SS的组成和结构与其他革兰氏阴性菌十分相似,其中编码分泌、转位和调控蛋白的基因位于基因组的5个操纵子上,编码效应蛋白或分子伴侣的基因分布在基因组的不同位点[4]。根据其功能可分为分泌装置(Secretion apparatus)或针管状复合体(Needle complex)、转位装置(Translocation apparatus)、效应蛋白或毒素(Effector toxins)及分子伴侣(Chaperones)四部分[5](图 1)。

|
| 图 1 铜绿假单胞菌T3SS结构示意图 Figure 1 Structure of the P. aeruginosa T3SS |
|
|
铜绿假单胞菌T3SS的分泌装置包括一个横跨细菌细胞内外膜的基础小体和一个指向胞外的针管状结构[6](图 1)。其中基础小体是一个双层的环状结构并通过中间的杆蛋白连接,而针管状结构是一种突出于菌体细胞膜表面的中空管状结构,由PscF蛋白(Type Ⅲ export protein,PscF)亚基组成,针状管道长约60 nm-120 nm,宽约6 nm-10 nm[3, 7]。当菌体侵染宿主细胞时,菌体产生的毒力因子先通过基础小体从细胞内部运送到针管状结构中,之后再通过针管状结构将效应蛋白送至转位装置[6]。此外,针管状结构还被认为是与宿主细胞接触的感受器[7]。
1.2 转位装置铜绿假单胞菌T3SS的转位装置是一个蛋白复合体,由其自身分泌的3种转位蛋白PopB、PopD及PcrV来行使转运功能[8](图 1)。其中PopB和PopD可与宿主细胞膜相互作用形成直径约2.8 nm-6.0 nm的转位孔道[9]。转位孔道的形成过程需要PcrV蛋白存在,但其本身并不参与组成转位孔道[10]。在整个效应蛋白转运过程中,铜绿假单胞菌首先与宿主细胞接触,激活T3SS的分泌装置并形成转位孔道,之后转位孔道插入宿主细胞的细胞膜上,靶向地向宿主细胞输入效应蛋白[11]。另外,研究表明T3SS的转运效率很高,超过99%的效应蛋白都可被注入到宿主细胞内[12]。
1.3 效应蛋白铜绿假单胞菌T3SS的效应蛋白主要有4种:胞外酶S (ExoS)、胞外酶T (ExoT)、胞外酶U (ExoU)和胞外酶Y (ExoY) (图 1),但并非所有菌株都产生这4种效应蛋白,而且ExoS与ExoU一般不在同一株菌中共存[13]。另有研究发现,铜绿假单胞菌中还存在两种新的T3SS效应蛋白,分别是PemA和PemB,其具体功能还有待进一步研究[14]。
铜绿假单胞菌PAO1菌株中的ExoS蛋白由453个氨基酸组成,是具有ADP核糖基转移酶(ADPRT)活性和GTP酶激活蛋白(GAP)活性的双重功能毒素[15]。其中GAP结构域可作用于真核细胞的小分子GTP酶Rho、Rac及Cdc42,从而导致肌动蛋白细胞骨架的破坏[16-17];而ADPRT活性可介导多种宿主蛋白(如Ras超家族蛋白)的ADP核糖基化,引起宿主细胞的信号传导过程失偶联,导致细胞变圆以及细胞凋亡[18-19]。
ExoT在结构和功能上与ExoS类似,由457个氨基酸组成,同样具有ADPRT和GAP结构域[20]。其中GAP结构域的作用与ExoS相似,均可使宿主细胞的Rho等GTP酶失活,进而破坏细胞骨架,导致细胞变圆、死亡,并能抑制宿主细胞的迁移及吞噬作用[21-22];虽然ExoS和ExoT的ADPRT活性均需要真核辅因子FAS的激活,二者的作用却存在一定的差异,ExoT的ADPRT结构域主要作用于真核细胞的Crk-Ⅰ和Crk-Ⅱ蛋白[23-24],ExoT也具有诱导细胞凋亡的能力,但其作用要晚于ExoS[25]。
ExoY是一种由378个氨基酸组成的腺苷酸环化酶,其活性需要一种未知的真核辅因子激活[26]。ExoY可引起宿主细胞内cAMP水平的升高,破坏肌动蛋白细胞骨架并导致血管内皮细胞通透性增加[27-28]。
ExoU是4种效应蛋白中毒性最强的一种,具有磷脂酶A2活性[29]。铜绿假单胞菌UCBPP-PA14菌株中的ExoU蛋白由687个氨基酸组成,其中142位的苏氨酸和344位的天冬氨酸残基对其酶活性具有关键作用[30]。当ExoU被注入到宿主细胞中时,会对细胞膜造成不可逆的损伤并引发细胞快速坏死,产生大量的炎症因子,最终导致严重的急性感染[31-32]。
1.4 分子伴侣T3SS的效应蛋白常常会和一些分子伴侣特异性地结合来维持蛋白质的稳定性,这些分子伴侣通常是一些分子量较小的酸性蛋白质,但分子伴侣本身并不被T3SS分泌[33]。在铜绿假单胞菌中,效应蛋白ExoS和ExoT的分子伴侣是SpcS蛋白[34],ExoU的分子伴侣为SpcU[35],而ExoY的分子伴侣到目前仍未发现。T3SS的某些分泌装置和转位装置蛋白同样也具有分子伴侣[3]。
2 铜绿假单胞菌T3SS的调控铜绿假单胞菌T3SS基因的表达与其所处的环境条件有着密不可分的关系。当细菌处于低Ca2+浓度的环境或与宿主细胞接触时,可以诱导T3SS的基因表达;而代谢压力、DNA损伤等一些不利因素均会抑制T3SS的活性[36-37]。针对不同环境条件的变化调整T3SS的表达,不仅有利于细菌保存充足的能量,还能减少抗体的产生,逃避宿主的免疫清除作用。近年来研究表明,铜绿假单胞菌中存在多个复杂的调控系统来控制T3SS的转录、翻译和分泌,这些大量的影响因素会使研究者更加清晰和完善地建立T3SS的调控网络,了解铜绿假单胞菌与环境和宿主之间的关系。
2.1 ExsA对T3SS的调控铜绿假单胞菌T3SS基因的表达受到转录激活子ExsA的直接调控[38]。ExsA属于AraC家族成员之一,该蛋白可以结合在T3SS基因的启动子区域(结合位点序列为TNAAAANA)来诱导T3SS的表达[39]。另有3个辅助蛋白ExsC、ExsE和ExsD可反馈调控ExsA的活性,将T3SS的转录与分泌过程偶联起来。在非分泌条件下,分泌蛋白ExsE存在于胞内,与ExsC结合,ExsD被ExsC释放并结合到ExsA上,从而抑制ExsA的活性[40-41]。在分泌条件下(低Ca2+浓度或与宿主细胞接触),ExsE被T3SS分泌到胞外,导致大量的ExsC与ExsD结合,从而释放出ExsA,激活T3SS的表达[42]。上述几种蛋白的编码基因均位于T3SS基因簇内部,属于T3SS内在调控因子。
2.2 cAMP/Vfr途径对T3SS的调控cAMP/Vfr途径对铜绿假单胞菌的群体感应系统(Quorum sensing,QS)、外毒素A的产生以及Ⅳ型菌毛介导的蹭行运动均具有调节作用,是该菌中非常重要的转录调控系统[43-45]。在铜绿假单胞菌中,腺苷酸环化酶CyaB对cAMP的合成起主要作用,合成的cAMP可通过与受体蛋白Vfr结合来影响基因的转录[46]。研究表明,外界环境信号也可以通过影响cAMP/Vfr途径来调控T3SS的表达。在低Ca2+浓度或高渗条件下,cyaB及vfr基因的转录水平增强,胞内cAMP的水平升高,T3SS的表达也相应增强,而vfr及cyaB基因的突变可导致T3SS的转录和分泌以及细菌细胞毒性明显降低[46-47]。有研究结果显示,Vfr蛋白可直接结合在exsA基因的启动子区域,激活exsA的转录从而调控T3SS的表达[48]。
2.3 Gac/Rsm系统与T3SS的关系在铜绿假单胞菌中,Gac/Rsm系统包括GacS/ GacA双组分系统,2个小RNA (sRNA) RsmY和RsmZ以及转录后调节蛋白RsmA。其中GacS为感应激酶,GacA为反应调节子。在一定条件下,GacS可通过自身磷酸化将磷酸基团传递给GacA,被激活的GacA可直接控制RsmY和RsmZ的表达从而拮抗RsmA的功能[49]。而RNA结合蛋白RsmA可正调控T3SS及细菌的运动能力等急性感染相关致病因子的表达,并负调控生物被膜形成及Ⅵ型分泌系统(Type Ⅵ secretion system,T6SS)等慢性感染相关基因的表达,从而影响铜绿假单胞菌的急慢性感染转变过程[50-52]。
在Gac/Rsm系统上游还存在3个双组分系统杂合感应激酶LadS、RetS和PA1611。其中LadS可通过一个复杂的磷酸化级联反应来激活Gac系统,同时,LadS激酶还可作为Ca2+受体蛋白来感应宿主信号,调控细菌从急性感染状态到慢性感染状态的转变过程[53-55]。RetS对Gac/Rsm系统的影响并不是通过经典的磷酸化途径,而是通过直接与GacS结合形成异源二聚体,使GacS不能磷酸化GacA,进而来影响下游基因的表达[56-57]。我们课题组在前期研究中发现了一个感应激酶PA1611,当PA1611蛋白高表达时,可显著抑制T3SS的表达并促进生物被膜的形成,细菌双杂交结果显示,PA1611可通过与RetS直接结合并释放GacS蛋白,从而激活Gac系统[58]。通过定点突变等实验我们确定了PA1611和RetS之间参与蛋白相互作用的关键氨基酸残基[59]。我们课题组还发现,与retS相邻的基因tspR (PA4857)突变之后可导致T3SS的表达降低,同时rsmY和rsmZ的表达明显升高,而tspR基因的表达又受到RetS的调控[60]。
2.4 c-di-GMP信号途径与T3SS的关系环二鸟苷酸(c-di-GMP)是细菌中普遍存在的第二信使分子,参与调控细菌的多种生物学功能,尤其在生物被膜的形成以及慢性感染的发生等方面起着非常重要的调节作用[61]。
有研究表明,在铜绿假单胞菌中,Gac/Rsm系统与c-di-GMP信号途径之间存在相互作用关系,retS的突变可导致c-di-GMP信号分子的水平显著升高,这两种途径可协同调控T3SS、T6SS以及生物被膜的形成[62]。而二鸟苷酸环化酶SadC被认为是将这两种调控途径联系起来的关键蛋白[63]。另有研究显示,suhB的缺失可在转录后水平抑制ExsA的表达,从而导致T3SS的表达和细菌毒性的丧失[64]。研究还显示,GacA的表达以及c-di-GMP的水平在suhB突变体中也有显著升高,SuhB可能通过Gac/Rsm以及c-di-GMP信号系统来影响T3SS的表达、细菌的运动性以及生物被膜的形成[65]。我们课题组发现,膜蛋白CmpX可通过调控PA1611基因的表达及细胞内c-di-GMP的水平来影响铜绿假单胞菌的急慢性感染过程[66]。此外,过表达二鸟苷酸环化酶DgcP可以抑制T3SS的表达[67]。过表达磷酸二酯酶FcsR同样可以抑制T3SS蛋白的表达和分泌,但这一抑制作用并不依赖FcsR的磷酸二酯酶活性[68]。参与c-di-GMP降解过程的寡核糖核酸酶Orn缺失之后也可导致T3SS表达及细菌致病性的显著降低[69]。
2.5 群体感应系统对T3SS的调控群体感应系统是一个与细菌群体密度相关的细胞间信号传导系统,它通过分泌并感应自诱导物信号分子来调控多种基因的表达[70]。铜绿假单胞菌中的QS系统分为3类:酰基高丝氨酸内酯分子介导的Las和Rhl系统以及喹诺酮类信号分子介导的PQS系统[71]。以往的研究表明,Rhl系统对T3SS相关基因的表达具有抑制作用,Las系统对T3SS没有调节作用[72-73]。稳定期σ因子RpoS也可以参与对T3SS的调控,RpoS还可以调控Rhl系统的表达,推测RpoS可能通过Rhl系统来影响T3SS的表达[73]。本课题组研究发现,PQS系统中的调控蛋白PqsR对T3SS效应蛋白基因的表达具有负调控作用[74]。PQS系统还可以影响T3SS效应蛋白的分泌[75]。另外,参与调控QS系统的转录调节子VqsM可直接结合在exsA的启动子区域来正调控T3SS的表达[76]。对PQS系统具有调节作用的PmpR蛋白也参与间接调控T3SS[77]。
2.6 T3SS与囊性纤维化的关系铜绿假单胞菌是造成囊性纤维化病人慢性呼吸道感染的最常见病原菌。临床上从CF病人呼吸道分离的菌株常常会过量表达藻酸盐并产生黏液型表型,这些黏液型菌株通常不再表达T3SS基因[78-79]。研究显示,藻酸盐的大量产生是mucA基因的突变所导致[80],MucA蛋白可抑制σ因子AlgU (又称AlgT)的活性[81],AlgU可激活包括双组分系统algZ/algR在内的多个藻酸盐合成相关基因的表达[82]。双组分系统中的AlgR蛋白又可以抑制vfr基因的转录并激活rsmY和rsmZ的表达[83-84],因此MucA可通过cAMP/Vfr以及Gac/Rsm途径来间接影响T3SS的表达。
2.7 环境压力对T3SS的影响除低Ca2+浓度以及与宿主细胞接触对T3SS的表达存在影响外,渗透压、DNA损伤、高Cu2+浓度、Mg2+转运、厌氧条件、温度以及代谢压力等均可以调控T3SS的表达。
研究显示,在培养基中添加高浓度的NaCl或蔗糖,可以激活T3SS基因的转录[47, 85]。同时在以上高渗透压条件下,细胞内cAMP的含量也会显著升高,由此推测高渗条件可能通过cAMP/Vfr途径来影响T3SS的表达[47]。
DNA损伤可作为一种调控基因表达的信号。在铜绿假单胞菌中,当DNA受到损伤首先会激活SOS系统修复受损DNA,在此应激反应条件下,转录抑制子PrtR被降解从而诱导产生PtrB蛋白,后者可以抑制T3SS的转录,这种抑制作用有助于细菌保存能量并及时修复DNA的损伤[86]。
研究表明,病人的烧伤组织可释放大量的Cu2+,而高Cu2+信号可以通过CopR-CopS双组分系统激活ptrA基因的转录,PtrA可直接与ExsA蛋白结合来抑制T3SS基因的转录[87-88]。Mg2+转运蛋白MgtE被发现可通过激活rsmY及rsmZ的转录来抑制T3SS基因的表达[89-90]。
参与厌氧条件下生长的一些酶及调节蛋白被发现也可以激活T3SS的表达[91-93],其中厌氧反应调节子Anr可通过双组分调节蛋白NarL抑制rsmY及rsmZ的转录来间接发挥作用[92]。此外,转录组分析结果显示,与环境温度(22 ℃)培养条件相比,接近宿主温度(37 ℃)的培养条件可以上调T3SS的表达[94]。
代谢压力对T3SS基因的表达也存在着影响,通常认为是由于菌体过度产生或消耗某种特定的代谢产物而引起。研究显示,转录调节子PsrA参与调节脂肪酸的降解过程,而长链脂肪酸可抑制PsrA蛋白对T3SS转录的激活[95-96]。丙氨酸脱氢酶合成基因(aceA和aceB)、葡萄糖转运调节基因(gtrS和gltR)以及亚精胺转运基因的突变可抑制T3SS的表达[46, 97-99]。过表达多药外排泵以及组氨酸转运和代谢相关基因同样可以抑制T3SS的表达[100-101]。代谢调控系统CbrA/CbrB/Crc也参与影响T3SS的表达[101-103]。此外,色氨酸代谢产物可参与细菌从低细胞密度到高细胞密度转变过程中T3SS表达的抑制[104]。细胞内的代谢失衡可能是引起以上这些影响的原因。有研究表明,乙酰辅酶A及其代谢衍生物可能是控制T3SS表达的关键因子[47]。
2.8 影响T3SS表达的其他因子除以上调控因素以外,在铜绿假单胞菌中还存在其他一些影响T3SS表达的因子。其中铜绿假单胞菌pfm基因突变之后对T3SS效应蛋白的水平会产生影响[105]。GTP酶TypA缺失之后可导致T3SS表达和毒力的降低[106]。LysR家族转录调节子BvlR可以抑制T3SS的表达[107]。RNA解旋酶DeaD可通过激活ExsA的翻译来启动T3SS的表达[108]。参与类脂A合成的2-羟基-十二烷酰转移酶PA0011可以抑制T3SS的转录和分泌[109]。多核苷酸磷酸化酶Pnp可通过影响RsmY和RsmZ的稳定性来调控T3SS的表达[110]。抗毒素系统HigB/HigA以及β-内酰胺酶AmpC也可以影响T3SS的表达[111-112]。DNA结合蛋白Fis可通过影响exsA的转录来调控T3SS的表达[113]。小RNA (Sr0161)可通过与exsA的mRNA结合来影响T3SS的表达[114]。
我们课题组以铜绿假单胞菌中T3SS效应蛋白基因exoS的表达为线索,通过转座突变的方法也筛选到了多个新的T3SS表达调控因子,其中包括PA1611等调控子[58]。此外有关ATP结合盒(ATP binding cassette,ABC)蛋白家族中的可溶性ATP酶PA4595对T3SS的调控机制也在进一步的研究中。
3 铜绿假单胞菌T3SS的针对性治疗策略目前针对铜绿假单胞菌感染的主要治疗手段是使用抗生素。但随着抗生素的应用,细菌的耐药性问题也日益加重。T3SS系统在铜绿假单胞菌致病性方面起着重要作用,因此开发以T3SS为靶点的治疗策略是目前研究的一个热点。近年来已有十几类小分子抑制剂或抗体疫苗被发现可以抑制T3SS的表达或功能,有些药物已经在临床试验验证当中[115]。
3.1 以T3SS转录调控为靶点从植物酚类化合物(Plant phenolic compounds)文库中筛选出来的TS027和TS103可以通过靶向Gac/Rsm系统来抑制T3SS的表达,这两类小分子化合物可通过诱导rsmY和rsmZ的表达来抑制exsA的转录,目前有关它们对T3SS介导的细胞毒性的作用仍在研究[116]。此外,抑制剂TS108和TS165可能通过exsA基因来影响exoS的基因表达及蛋白产量[117]。
N-羟基苯并咪唑(N-hydroxybenzimidazoles)可以与ExsA蛋白相互作用,阻止ExsA结合到DNA的启动子区域[118]。一般认为N-羟基苯并咪唑的结合口袋位于ExsA的DNA结合结构域中,该口袋中特定氨基酸的取代可以导致N-羟基苯并咪唑敏感性的改变,从而帮助人们确定配体的结合位点并设计更高亲和力的抑制剂[119]。
水杨酰肼(Salicylidene acylhydrazides)已被筛选为不同细菌物种的T3SS转录抑制剂。在铜绿假单胞菌中,也筛选出了多个该类型的T3SS抑制剂,其中包括INP0341,但其抗假单胞菌的作用机制目前尚未阐明[120]。
3.2 以T3SS装置功能为靶点T3SS的分泌和转位装置具有复杂的结构,破坏其中某一蛋白的功能就会威胁整个系统的活性,这为多种药物干预提供了可能。
羟基喹啉(Hydroxyquinolines)类化合物INP1855可以作用于T3SS以及鞭毛装置,抑制ExoS的分泌以及鞭毛运动,一些实验证实此类化合物在保护真核细胞免受T3SS毒害作用等方面具有显著效果[121]。噻唑烷酮(Thiazolidinones)类化合物可以抑制铜绿假单胞菌中T3SS和Ⅱ型分泌系统(Type Ⅱ secretion system,T2SS)的分泌活性[122]。苯氧基酰胺(Phenoxyacetamides)类化合物MBX1641可降低T3SS的分泌和转位活性,而不影响T2SS的活性和其他蛋白的表达[123]。有研究者在MBX1641结构的基础上合成了一系列新的衍生物,部分化合物对exoS基因的表达有明显的抑制作用,其抑制活性强于MBX1641[124]。此类抑制剂除可以抑制T3SS表达外,还可能通过影响其它毒性因子来影响铜绿假单胞菌的致病性[125]。
T3SS转位装置常被作为抗体类抑制剂的作用靶点。已有两类抗PcrV抗体被显示可抑制效应蛋白转位并恢复巨噬细胞的吞噬功能,动物感染和免疫治疗实验也证实了该类抗体的有效性[126-128]。尽管菌株中的PcrV序列多变,抗PcrV抗体仍可有效地降低众多临床分离株的细胞毒性,这也表明了它们在临床上的适用性[129]。目前部分抗PcrV疫苗已完成Ⅰ期或Ⅱ期临床试验[130-131]。
3.3 以T3SS效应蛋白为靶点Exosin可通过作为NAD+底物的竞争性抑制剂来损害ExoS的ADP核糖基转移酶活性,从而降低ExoS对哺乳动物细胞的细胞毒性[132]。Pseudolipasin A可以特异性地抑制ExoU的磷脂酶A2活性,而不影响被测试的其他真核磷脂酶,这一现象证实该物质可以保护宿主细胞免受ExoU表达菌株的细胞毒性[133]。此外芳基磺酰胺(Arylsulfonamides)类化合物也可以抑制ExoU介导的细胞毒性,但其作用与Pseudolipasin A相比较弱[134]。
4 总结与展望铜绿假单胞菌是主要的医院里感染条件致病菌,拥有多种致病机制和耐药机制,其中T3SS是该菌致病过程的关键毒性因子。本文针对T3SS的复杂调控机制和针对性治疗策略进行了着重讨论,概括了影响T3SS表达的多种环境和宿主信号以及直接或间接的T3SS调控途径。在众多的调控途径中,其中一些调节子只调控T3SS,如ExsA属于T3SS内在调控因子;另有一些调控途径可以影响包括T3SS在内的多种致病相关基因的表达,如Gac/Rsm系统以及c-di-GMP信号途径等可通过其调控作用影响铜绿假单胞菌的急慢性感染转变过程。以上有关调控机制的研究也为T3SS针对性治疗策略的开发提供了理论基础。
尽管近些年有关铜绿假单胞菌中T3SS的研究取得了很多进展,但这中间仍存在许多亟待解决的问题。今后的研究方向应是进一步探索T3SS的致病机理和调控机制,继续筛选与T3SS调控相关的关键基因,并将各种环境及宿主信号与胞内的复杂调控途径联系起来,构建出一个完整而全面的调控网络系统。同时,还应致力于以T3SS为靶点的药物研发,找到控制铜绿假单胞菌感染的更优方案。目前有关T3SS小分子抑制剂的研究尚处于药物筛选和实验室研究阶段,仅有部分抗体疫苗在进行临床试验。通过结合生物信息学和结构生物学等研究手段,进一步筛选更有效的T3SS抑制剂,明确药物的作用原理,揭示病原菌与宿主相互作用的新机制,有关这一领域的深入研究将有助于推动抗致病性药物的临床应用进程。
| [1] |
Lyczak JB, Cannon CL, Pier GB. Lung infections associated with cystic fibrosis[J]. Clinical Microbiology Reviews, 2002, 15(2): 194-222. DOI:10.1128/CMR.15.2.194-222.2002 |
| [2] |
Trautmann M, Lepper PM, Haller M. Ecology of Pseudomonas aeruginosa in the intensive care unit and the evolving role of water outlets as a reservoir of the organism[J]. American Journal of Infection Control, 2005, 33(5S): S41-S49. |
| [3] |
Hauser AR. The type Ⅲ secretion system of Pseudomonas aeruginosa: infection by injection[J]. Nature Reviews Microbiology, 2009, 7(9): 654-665. DOI:10.1038/nrmicro2199 |
| [4] |
Yahr TL, Wolfgang MC. Transcriptional regulation of the Pseudomonas aeruginosa type Ⅲ secretion system[J]. Molecular Microbiology, 2006, 62(3): 631-640. |
| [5] |
Galle M, Carpentier I, Beyaert R. Structure and function of the type Ⅲ secretion system of Pseudomonas aeruginosa[J]. Current Protein & Peptide Science, 2012, 13(8): 831-842. |
| [6] |
Cornelis GR. The type Ⅲ secretion injectisome[J]. Nature Reviews Microbiology, 2006, 4(11): 811-825. DOI:10.1038/nrmicro1526 |
| [7] |
Pastor A, Chabert J, Louwagie M, et al. PscF is a major component of the Pseudomonas aeruginosa type Ⅲ secretion needle[J]. FEMS Microbiology Letters, 2005, 253(1): 95-101. DOI:10.1016/j.femsle.2005.09.028 |
| [8] |
Mueller CA, Broz P, Cornelis GR. The type Ⅲ secretion system tip complex and translocon[J]. Molecular Microbiology, 2008, 68(5): 1085-1095. DOI:10.1111/j.1365-2958.2008.06237.x |
| [9] |
Schoehn G, Di Guilmi AM, Lemaire D, et al. Oligomerization of type Ⅲ secretion proteins PopB and PopD precedes pore formation in Pseudomonas[J]. The EMBO Journal, 2003, 22(19): 4957-4967. DOI:10.1093/emboj/cdg499 |
| [10] |
Goure J, Pastor A, Faudry E, et al. The V antigen of Pseudomonas aeruginosa is required for assembly of the functional PopB/PopD translocation pore in host cell membranes[J]. Infection and Immunity, 2004, 72(8): 4741-4750. DOI:10.1128/IAI.72.8.4741-4750.2004 |
| [11] |
Faudry E, Vernier G, Neumann E, et al. Synergistic pore formation by type Ⅲ toxin translocators of Pseudomonas aeruginosa[J]. Biochemistry, 2006, 45(26): 8117-8123. DOI:10.1021/bi060452+ |
| [12] |
Sundin C, Thelaus J, BrÖms JE, et al. Polarisation of type Ⅲ translocation by Pseudomonas aeruginosa requires PcrG, PcrV and PopN[J]. Microbial Pathogenesis, 2004, 37(6): 313-322. DOI:10.1016/j.micpath.2004.10.005 |
| [13] |
Feltman H, Schulert G, Khan S, et al. Prevalence of type Ⅲ secretion genes in clinical and environmental isolates of Pseudomonas aeruginosa[J]. Microbiology, 2001, 147(10): 2659-2669. DOI:10.1099/00221287-147-10-2659 |
| [14] |
Burstein D, Satanower S, Simovitch M, et al. Novel type Ⅲ effectors in Pseudomonas aeruginosa[J]. mBio, 2015, 6(2): e00161-15. |
| [15] |
Rocha CL, Coburn J, Rucks EA, et al. Characterization of Pseudomonas aeruginosa exoenzyme S as a bifunctional enzyme in J774A.1 macrophages[J]. Infection and Immunity, 2003, 71(9): 5296-5305. DOI:10.1128/IAI.71.9.5296-5305.2003 |
| [16] |
Pederson KJ, Vallis AJ, Aktories K, et al. The amino-terminal domain of Pseudomonas aeruginosa ExoS disrupts actin filaments via small-molecular-weight GTP-binding proteins[J]. Molecular Microbiology, 1999, 32(2): 393-401. |
| [17] |
Würtele M, Wolf E, Pederson KJ, et al. How the Pseudomonas aeruginosa ExoS toxin downregulates Rac[J]. Nature Structural Biology, 2001, 8(1): 23-26. DOI:10.1038/83007 |
| [18] |
Ganesan AK, Frank DW, Misra RP, et al. Pseudomonas aeruginosa exoenzyme S ADP-ribosylates Ras at multiple sites[J]. Journal of Biological Chemistry, 1998, 273(13): 7332-7337. DOI:10.1074/jbc.273.13.7332 |
| [19] |
Kaufman MR, Jia JH, Zeng L, et al. Pseudomonas aeruginosa mediated apoptosis requires the ADP-ribosylating activity of ExoS[J]. Microbiology, 2000, 146(10): 2531-2541. DOI:10.1099/00221287-146-10-2531 |
| [20] |
Barbieri JT, Sun J. Pseudomonas aeruginosa ExoS and ExoT[A]//Amara SG, Bamberg E, Gudermann T, et al. Reviews of Physiology, Biochemistry and Pharmacology[M]. Berlin, Heidelberg: Springer, 2005: 79-92
|
| [21] |
Kazmierczak BI, Engel JN. Pseudomonas aeruginosa ExoT acts in vivo as a GTPase-activating protein for RhoA, Rac1, and Cdc42[J]. Infection and Immunity, 2002, 70(4): 2198-2205. DOI:10.1128/IAI.70.4.2198-2205.2002 |
| [22] |
Krall R, Schmidt G, Aktories K, et al. Pseudomonas aeruginosa ExoT is a Rho GTPase-activating protein[J]. Infection and Immunity, 2000, 68(10): 6066-6068. DOI:10.1128/IAI.68.10.6066-6068.2000 |
| [23] |
Deng Q, Sun JJ, Barbieri JT. Uncoupling Crk signal transduction by Pseudomonas exoenzyme T[J]. Journal of Biological Chemistry, 2005, 280(43): 35953-35960. DOI:10.1074/jbc.M504901200 |
| [24] |
Sun JJ, Barbieri JT. Pseudomonas aeruginosa ExoT ADP-ribosylates CT10 regulator of kinase (Crk) proteins[J]. Journal of Biological Chemistry, 2003, 278(35): 32794-32800. DOI:10.1074/jbc.M304290200 |
| [25] |
Shafikhani SH, Morales C, Engel J. The Pseudomonas aeruginosa type Ⅲ secreted toxin ExoT is necessary and sufficient to induce apoptosis in epithelial cells[J]. Cellular Microbiology, 2008, 10(4): 994-1007. DOI:10.1111/j.1462-5822.2007.01102.x |
| [26] |
Yahr TL, Vallis AJ, Hancock MK, et al. ExoY, an adenylate cyclase secreted by the Pseudomonas aeruginosa type Ⅲ system[J]. Proceedings of the National Academy of Sciences of the United States of America, 1998, 95(23): 13899-13904. DOI:10.1073/pnas.95.23.13899 |
| [27] |
Cowell BA, Evans DJ, Fleiszig SMJ. Actin cytoskeleton disruption by ExoY and its effects on Pseudomonas aeruginosa invasion[J]. FEMS Microbiology Letters, 2005, 250(1): 71-76. DOI:10.1016/j.femsle.2005.06.044 |
| [28] |
Sayner SL, Frank DW, King J, et al. Paradoxical cAMP-induced lung endothelial hyperpermeability revealed by Pseudomonas aeruginosa ExoY[J]. Circulation Research, 2004, 95(2): 196-203. DOI:10.1161/01.RES.0000134922.25721.d9 |
| [29] |
Sato H, Feix JB, Hillard CJ, et al. Characterization of phospholipase activity of the Pseudomonas aeruginosa type Ⅲ cytotoxin, ExoU[J]. Journal of Bacteriology, 2005, 187(3): 1192-1195. DOI:10.1128/JB.187.3.1192-1195.2005 |
| [30] |
Rabin SDP, Hauser AR. Functional regions of the Pseudomonas aeruginosa cytotoxin ExoU[J]. Infection and Immunity, 2005, 73(1): 573-582. DOI:10.1128/IAI.73.1.573-582.2005 |
| [31] |
Sato H, Frank DW, Hillard CJ, et al. The mechanism of action of the Pseudomonas aeruginosa-encoded type Ⅲ cytotoxin, ExoU[J]. The EMBO Journal, 2003, 22(12): 2959-2969. DOI:10.1093/emboj/cdg290 |
| [32] |
Schulert GS, Feltman H, Rabin SDP, et al. Secretion of the toxin ExoU is a marker for highly virulent Pseudomonas aeruginosa isolates obtained from patients with hospital-acquired pneumonia[J]. The Journal of Infectious Diseases, 2003, 188(11): 1695-1706. DOI:10.1086/jid.2003.188.issue-11 |
| [33] |
Parsot C, Hamiaux C, Page AL. The various and varying roles of specific chaperones in type Ⅲ secretion systems[J]. Current Opinion in Microbiology, 2003, 6(1): 7-14. |
| [34] |
Shen DK, Quenee L, Bonnet M, et al. Orf1/SpcS Chaperones ExoS for Type Three Secretion by Pseudomonas aeruginosa[J]. Biomedical and Environmental Sciences, 2008, 21(2): 103-109. DOI:10.1016/S0895-3988(08)60014-8 |
| [35] |
Finck-Barbancon V, Yahr TL, Frank DW. Identification and characterization of SpcU, a chaperone required for efficient secretion of the ExoU cytotoxin[J]. Journal of Bacteriology, 1998, 180(23): 6224-6231. |
| [36] |
Diaz MR, King JM, Yahr TL. Intrinsic and extrinsic regulation of type Ⅲ secretion gene expression in Pseudomonas aeruginosa[J]. Frontiers in Microbiology, 2011, 2: 89. |
| [37] |
Luo Q, Jin SG. Molecular determinants in regulating Pseudomonas aeruginosa type Ⅲ secretion system—a review[J]. Acta Microbiologica Sinica, 2008, 48(10): 1413-1417. (in Chinese) 罗勤, 金守光. 铜绿假单胞菌Ⅲ型分泌系统的分子调控机制[J]. 微生物学报, 2008, 48(10): 1413-1417. DOI:10.3321/j.issn:0001-6209.2008.10.025 |
| [38] |
Frank DW. The exoenzyme S regulon of Pseudomonas aeruginosa[J]. Molecular Microbiology, 1997, 26(4): 621-629. |
| [39] |
Hovey AK, Frank DW. Analyses of the DNA-binding and transcriptional activation properties of ExsA, the transcriptional activator of the Pseudomonas aeruginosa exoenzyme S regulon[J]. Journal of Bacteriology, 1995, 177(15): 4427-4436. DOI:10.1128/jb.177.15.4427-4436.1995 |
| [40] |
Dasgupta N, Lykken GL, Wolfgang MC, et al. A novel anti-anti-activator mechanism regulates expression of the Pseudomonas aeruginosa type Ⅲ secretion system[J]. Molecular Microbiology, 2004, 53(1): 297-308. DOI:10.1111/j.1365-2958.2004.04128.x |
| [41] |
McCaw ML, Lykken GL, Singh PK, et al. ExsD is a negative regulator of the Pseudomonas aeruginosa type Ⅲ secretion regulon[J]. Molecular Microbiology, 2002, 46(4): 1123-1133. |
| [42] |
Urbanowski ML, Brutinel ED, Yahr TL. Translocation of ExsE into Chinese hamster ovary cells is required for transcriptional induction of the Pseudomonas aeruginosa type Ⅲ secretion system[J]. Infection and Immunity, 2007, 75(9): 4432-4439. DOI:10.1128/IAI.00664-07 |
| [43] |
Albus AM, Pesci EC, Runyen-Janecky LJ, et al. Vfr controls quorum sensing in Pseudomonas aeruginosa[J]. Journal of Bacteriology, 1997, 179(12): 3928-3935. DOI:10.1128/jb.179.12.3928-3935.1997 |
| [44] |
Beatson SA, Whitchurch CB, Sargent JL, et al. Differential regulation of twitching motility and elastase production by Vfr in Pseudomonas aeruginosa[J]. Journal of Bacteriology, 2002, 184(13): 3605-3613. DOI:10.1128/JB.184.13.3605-3613.2002 |
| [45] |
West SE, Sample AK, Runyen-Janecky LJ. The vfr gene product, required for Pseudomonas aeruginosa exotoxin A and protease production, belongs to the cyclic AMP receptor protein family[J]. Journal of Bacteriology, 1994, 176(24): 7532-7542. DOI:10.1128/jb.176.24.7532-7542.1994 |
| [46] |
Wolfgang MC, Lee VT, Gilmore ME, et al. Coordinate regulation of bacterial virulence genes by a novel adenylate cyclase-dependent signaling pathway[J]. Developmental Cell, 2003, 4(2): 253-263. DOI:10.1016/S1534-5807(03)00019-4 |
| [47] |
Rietsch A, Mekalanos JJ. Metabolic regulation of type Ⅲ secretion gene expression in Pseudomonas aeruginosa[J]. Molecular Microbiology, 2006, 59(3): 807-820. |
| [48] |
Marsden AE, Intile PJ, Schulmeyer KH, et al. Vfr directly activates exsA transcription to regulate expression of the Pseudomonas aeruginosa type Ⅲ secretion system[J]. Journal of Bacteriology, 2016, 198(9): 1442-1450. DOI:10.1128/JB.00049-16 |
| [49] |
Brencic A, McFarland KA, McManus HR, et al. The GacS/GacA signal transduction system of Pseudomonas aeruginosa acts exclusively through its control over the transcription of the RsmY and RsmZ regulatory small RNAs[J]. Molecular Microbiology, 2009, 73(3): 434-445. |
| [50] |
Allsopp LP, Wood TE, Howard SA, et al. RsmA and AmrZ orchestrate the assembly of all three type Ⅵ secretion systems in Pseudomonas aeruginosa[J]. Proceedings of the National Academy of Sciences of the United States of America, 2017, 114(29): 7707-7712. DOI:10.1073/pnas.1700286114 |
| [51] |
Brencic A, Lory S. Determination of the regulon and identification of novel mRNA targets of Pseudomonas aeruginosa RsmA[J]. Molecular Microbiology, 2009, 72(3): 612-632. |
| [52] |
Irie Y, Starkey M, Edwards AN, et al. Pseudomonas aeruginosa biofilm matrix polysaccharide Psl is regulated transcriptionally by RpoS and post-transcriptionally by RsmA[J]. Molecular Microbiology, 2010, 78(1): 158-172. |
| [53] |
Broder UN, Jaeger T, Jenal U. LadS is a calcium-responsive kinase that induces acute-to-chronic virulence switch in Pseudomonas aeruginosa[J]. Nature Microbiology, 2016, 2: 16184. |
| [54] |
Chambonnier G, Roux L, Redelberger D, et al. The hybrid histidine kinase LadS forms a multicomponent signal transduction system with the GacS/GacA two-component system in Pseudomonas aeruginosa[J]. PLoS Genetics, 2016, 12(5): e1006032. DOI:10.1371/journal.pgen.1006032 |
| [55] |
Ventre I, Goodman AL, Vallet-Gely I, et al. Multiple sensors control reciprocal expression of Pseudomonas aeruginosa regulatory RNA and virulence genes[J]. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(1): 171-176. DOI:10.1073/pnas.0507407103 |
| [56] |
Goodman AL, Kulasekara B, Rietsch A, et al. A signaling network reciprocally regulates genes associated with acute infection and chronic persistence in Pseudomonas aeruginosa[J]. Developmental Cell, 2004, 7(5): 745-754. DOI:10.1016/j.devcel.2004.08.020 |
| [57] |
Goodman AL, Merighi M, Hyodo M, et al. Direct interaction between sensor kinase proteins mediates acute and chronic disease phenotypes in a bacterial pathogen[J]. Genes & Development, 2009, 23(2): 249-259. |
| [58] |
Kong WN, Chen L, Zhao JQ, et al. Hybrid sensor kinase PA1611 in Pseudomonas aeruginosa regulates transitions between acute and chronic infection through direct interaction with RetS[J]. Molecular Microbiology, 2013, 88(4): 784-797. |
| [59] |
Bhagirath AY, Pydi SP, Li YQ, et al. Characterization of the direct interaction between hybrid sensor kinases PA1611 and RetS that controls biofilm formation and the type Ⅲ secretion system in Pseudomonas aeruginosa[J]. ACS Infectious Diseases, 2017, 3(2): 162-175. DOI:10.1021/acsinfecdis.6b00153 |
| [60] |
Zhu M, Zhao JR, Kang HP, et al. Modulation of type Ⅲ secretion system in Pseudomonas aeruginosa: involvement of the PA4857 gene product[J]. Frontiers in Microbiology, 2016, 7: 7. |
| [61] |
RÖmling U, Galperin MY, Gomelsky M. Cyclic di-GMP: the first 25 years of a universal bacterial second messenger[J]. Microbiology and Molecular Biology Reviews, 2013, 77(1): 1-52. |
| [62] |
Moscoso JA, Mikkelsen H, Heeb S, et al. The Pseudomonas aeruginosa sensor RetS switches type Ⅲ and type Ⅵ secretion via c-di-GMP signalling[J]. Environmental Microbiology, 2011, 13(12): 3128-3138. DOI:10.1111/j.1462-2920.2011.02595.x |
| [63] |
Moscoso JA, Jaeger T, Valentini M, et al. The diguanylate cyclase SadC is a central player in Gac/Rsm-mediated biofilm formation in Pseudomonas aeruginosa[J]. Journal of Bacteriology, 2014, 196(23): 4081-4088. DOI:10.1128/JB.01850-14 |
| [64] |
Li KW, Xu C, Jin YX, et al. SuhB is a regulator of multiple virulence genes and essential for pathogenesis of Pseudomonas aeruginosa[J]. mBio, 2013, 4(6): e00419-13. |
| [65] |
Li KW, Yang GJ, Debru AB, et al. SuhB regulates the motile-sessile switch in Pseudomonas aeruginosa through the Gac/Rsm pathway and c-di-GMP signaling[J]. Frontiers in Microbiology, 2017, 8: 1045. DOI:10.3389/fmicb.2017.01045 |
| [66] |
Bhagirath AY, Somayajula D, Li YQ, et al. CmpX affects virulence in Pseudomonas aeruginosa through the Gac/Rsm signaling pathway and by modulating c-di-GMP levels[J]. The Journal of Membrane Biology, 2018, 251(1): 35-49. |
| [67] |
Aragon IM, Pérez-Mendoza D, Moscoso JA, et al. Diguanylate cyclase DgcP is involved in plant and human Pseudomonas spp. infections[J]. Environmental Microbiology, 2015, 17(11): 4332-4351. DOI:10.1111/1462-2920.12856 |
| [68] |
Rossello J, Lima A, Gil M, et al. The EAL-domain protein FcsR regulates flagella, chemotaxis and type Ⅲ secretion system in Pseudomonas aeruginosa by a phosphodiesterase independent mechanism[J]. Scientific Reports, 2017, 7(1): 10281. DOI:10.1038/s41598-017-09926-3 |
| [69] |
Chen GK, Zhao Q, Zhu F, et al. Oligoribonuclease is required for the type Ⅲ secretion system and pathogenesis of Pseudomonas aeruginosa[J]. Microbiological Research, 2016, 188-189: 90-96. DOI:10.1016/j.micres.2016.05.002 |
| [70] |
Abisado RG, Benomar S, Klaus JR, et al. Bacterial quorum sensing and microbial community interactions[J]. mBio, 2018, 9(3): e02331-17. |
| [71] |
Lee J, Zhang LH. The hierarchy quorum sensing network in Pseudomonas aeruginosa[J]. Protein & Cell, 2015, 6(1): 26-41. |
| [72] |
Bleves S, Soscia C, Nogueira-Orlandi P, et al. Quorum sensing negatively controls type Ⅲ secretion regulon expression in Pseudomonas aeruginosa PAO1[J]. Journal of Bacteriology, 2005, 187(11): 3898-3902. DOI:10.1128/JB.187.11.3898-3902.2005 |
| [73] |
Hogardt M, Roeder M, Schreff AM, et al. Expression of Pseudomonas aeruginosa exoS is controlled by quorum sensing and RpoS[J]. Microbiology, 2004, 150(4): 843-851. |
| [74] |
Kong WN, Liang HH, Shen LX, et al. Regulation of type Ⅲ secretion system by Rhl and PQS quorum sensing systems in Pseudomonas aeruginosa[J]. Acta Microbiologica Sinica, 2009, 49(9): 1158-1164. (in Chinese) 孔伟娜, 梁海华, 沈立新, 等. 铜绿假单胞菌中Ⅲ型分泌系统受Rhl和PQS群体感应系统调节[J]. 微生物学报, 2009, 49(9): 1158-1164. DOI:10.3321/j.issn:0001-6209.2009.09.005 |
| [75] |
Singh G, Wu B, Baek MS, et al. Secretion of Pseudomonas aeruginosa type Ⅲ cytotoxins is dependent on pseudomonas quinolone signal concentration[J]. Microbial Pathogenesis, 2010, 49(4): 196-203. DOI:10.1016/j.micpath.2010.05.013 |
| [76] |
Liang HH, Deng X, Li XF, et al. Molecular mechanisms of master regulator VqsM mediating quorum-sensing and antibiotic resistance in Pseudomonas aeruginosa[J]. Nucleic Acids Research, 2014, 42(16): 10307-10320. DOI:10.1093/nar/gku586 |
| [77] |
Liang HH, Kong WN, Shen T, et al. The effect of pmpR on the type Ⅲ secretion system in Pseudomonas aeruginosa[J]. Chinese Science Bulletin, 2012, 57(19): 2413-2418. DOI:10.1007/s11434-011-4941-x |
| [78] |
Allmond LR, Ajayi T, Moriyama K, et al. V-antigen genotype and phenotype analyses of clinical isolates of Pseudomonas aeruginosa[J]. Journal of Clinical Microbiology, 2004, 42(8): 3857-3860. DOI:10.1128/JCM.42.8.3857-3860.2004 |
| [79] |
Martin DW, Schurr MJ, Mudd MH, et al. Mechanism of conversion to mucoidy in Pseudomonas aeruginosa infecting cystic fibrosis patients[J]. Proceedings of the National Academy of Sciences of the United States of America, 1993, 90(18): 8377-8381. DOI:10.1073/pnas.90.18.8377 |
| [80] |
Wu WH, Badrane H, Arora S, et al. MucA-mediated coordination of type Ⅲ secretion and alginate synthesis in Pseudomonas aeruginosa[J]. Journal of Bacteriology, 2004, 186(22): 7575-7585. DOI:10.1128/JB.186.22.7575-7585.2004 |
| [81] |
Schurr MJ, Yu H, Martinez-Salazar JM, et al. Control of AlgU, a member of the sigma E-like family of stress sigma factors, by the negative regulators MucA and MucB and Pseudomonas aeruginosa conversion to mucoidy in cystic fibrosis[J]. Journal of Bacteriology, 1996, 178(16): 4997-5004. DOI:10.1128/jb.178.16.4997-5004.1996 |
| [82] |
Wozniak DJ, Ohman DE. Transcriptional analysis of the Pseudomonas aeruginosa genes algR, algB, and algD reveals a hierarchy of alginate gene expression which is modulated by algT[J]. Journal of Bacteriology, 1994, 176(19): 6007-6014. DOI:10.1128/jb.176.19.6007-6014.1994 |
| [83] |
Intile PJ, Diaz MR, Urbanowski ML, et al. The AlgZR two-component system recalibrates the RsmAYZ posttranscriptional regulatory system to inhibit expression of the Pseudomonas aeruginosa type Ⅲ secretion system[J]. Journal of Bacteriology, 2014, 196(2): 357-366. DOI:10.1128/JB.01199-13 |
| [84] |
Jones AK, Fulcher NB, Balzer GJ, et al. Activation of the Pseudomonas aeruginosa AlgU regulon through mucA mutation inhibits cyclic AMP/Vfr signaling[J]. Journal of Bacteriology, 2010, 192(21): 5709-5717. DOI:10.1128/JB.00526-10 |
| [85] |
Aspedon A, Palmer K, Whiteley M. Microarray analysis of the osmotic stress response in Pseudomonas aeruginosa[J]. Journal of Bacteriology, 2006, 188(7): 2721-2725. DOI:10.1128/JB.188.7.2721-2725.2006 |
| [86] |
Wu WH, Jin SG. PtrB of Pseudomonas aeruginosa suppresses the type Ⅲ secretion system under the stress of DNA damage[J]. Journal of Bacteriology, 2005, 187(17): 6058-6068. DOI:10.1128/JB.187.17.6058-6068.2005 |
| [87] |
Elsen S, Ragno M, Attree I. PtrA is a periplasmic protein involved in Cu tolerance in Pseudomonas aeruginosa[J]. Journal of Bacteriology, 2011, 193(13): 3376-3378. DOI:10.1128/JB.00159-11 |
| [88] |
Ha UH, Kim J, Badrane H, et al. An in vivo inducible gene of Pseudomonas aeruginosa encodes an anti-ExsA to suppress the type Ⅲ secretion system[J]. Molecular Microbiology, 2004, 54(2): 307-320. DOI:10.1111/j.1365-2958.2004.04282.x |
| [89] |
Anderson GG, Yahr TL, Lovewell RR, et al. The Pseudomonas aeruginosa magnesium transporter MgtE inhibits transcription of the type Ⅲ secretion system[J]. Infection and Immunity, 2010, 78(3): 1239-1249. DOI:10.1128/IAI.00865-09 |
| [90] |
Chakravarty S, Melton CN, Bailin A, et al. Pseudomonas aeruginosa magnesium transporter MgtE inhibits type Ⅲ secretion system gene expression by stimulating rsmYZ transcription[J]. Journal of Bacteriology, 2017, 199(23): e00268-17. |
| [91] |
Chung JCS, Rzhepishevska O, Ramstedt M, et al. Type Ⅲ secretion system expression in oxygen-limited Pseudomonas aeruginosa cultures is stimulated by isocitrate lyase activity[J]. Open Biology, 2013, 3(1): 120131. DOI:10.1098/rsob.120131 |
| [92] |
O'Callaghan J, Reen FJ, Adams C, et al. Low oxygen induces the type Ⅲ secretion system in Pseudomonas aeruginosa via modulation of the small RNAs rsmZ and rsmY[J]. Microbiology, 2011, 157(12): 3417-3428. DOI:10.1099/mic.0.052050-0 |
| [93] |
Van Alst NE, Wellington M, Clark VL, et al. Nitrite reductase NirS is required for type Ⅲ secretion system expression and virulence in the human monocyte cell line THP-1 by Pseudomonas aeruginosa[J]. Infection and Immunity, 2009, 77(10): 4446-4454. DOI:10.1128/IAI.00822-09 |
| [94] |
Barbier M, Damron FH, Bielecki P, et al. From the environment to the host: re-wiring of the transcriptome of Pseudomonas aeruginosa from 22 ℃ to 37 ℃[J]. PLoS One, 2014, 9(2): e89941. DOI:10.1371/journal.pone.0089941 |
| [95] |
Kang Y, Lunin VV, Skarina T, et al. The long-chain fatty acid sensor, PsrA, modulates the expression of rpoS and the type Ⅲ secretion exsCEBA operon in Pseudomonas aeruginosa[J]. Molecular Microbiology, 2009, 73(1): 120-136. DOI:10.1111/mmi.2009.73.issue-1 |
| [96] |
Shen DK, Filopon D, Kuhn L, et al. PsrA is a positive transcriptional regulator of the type Ⅲ secretion system in Pseudomonas aeruginosa[J]. Infection and Immunity, 2006, 74(2): 1121-1129. DOI:10.1128/IAI.74.2.1121-1129.2006 |
| [97] |
Dacheux D, Epaulard O, de Groot A, et al. Activation of the Pseudomonas aeruginosa type Ⅲ secretion system requires an intact pyruvate dehydrogenase aceAB operon[J]. Infection and Immunity, 2002, 70(7): 3973-3977. DOI:10.1128/IAI.70.7.3973-3977.2002 |
| [98] |
O'Callaghan J, Reen JF, Adams C, et al. A novel host-responsive sensor mediates virulence and type Ⅲ secretion during Pseudomonas aeruginosa-host cell interactions[J]. Microbiology, 2012, 158(Pt 4): 1057-1070. |
| [99] |
Zhou L, Wang J, Zhang LH. Modulation of bacterial type Ⅲ secretion system by a spermidine transporter dependent signaling pathway[J]. PLoS One, 2007, 2(12): e1291. DOI:10.1371/journal.pone.0001291 |
| [100] |
Linares JF, López JA, Camafeita E, et al. Overexpression of the multidrug efflux pumps MexCD-OprJ and MexEF-OprN is associated with a reduction of type Ⅲ secretion in Pseudomonas aeruginosa[J]. Journal of Bacteriology, 2005, 187(4): 1384-1391. DOI:10.1128/JB.187.4.1384-1391.2005 |
| [101] |
Rietsch A, Wolfgang MC, Mekalanos JJ. Effect of metabolic imbalance on expression of type Ⅲ secretion genes in Pseudomonas aeruginosa[J]. Infection and Immunity, 2004, 72(3): 1383-1390. DOI:10.1128/IAI.72.3.1383-1390.2004 |
| [102] |
Dong YH, Zhang XF, Zhang LH. The global regulator Crc plays a multifaceted role in modulation of type Ⅲ secretion system in Pseudomonas aeruginosa[J]. MicrobiologyOpen, 2013, 2(1): 161-172. DOI:10.1002/mbo3.54 |
| [103] |
Yeung ATY, Bains M, Hancock REW. The sensor kinase CbrA is a global regulator that modulates metabolism, virulence, and antibiotic resistance in Pseudomonas aeruginosa[J]. Journal of Bacteriology, 2011, 193(4): 918-931. DOI:10.1128/JB.00911-10 |
| [104] |
Shen DK, Filopon D, Chaker H, et al. High-cell-density regulation of the Pseudomonas aeruginosa type Ⅲ secretion system: implications for tryptophan catabolites[J]. Microbiology, 2008, 154(8): 2195-2208. DOI:10.1099/mic.0.2007/013680-0 |
| [105] |
Mou R, Du X, Wang XH, et al. Pseudomonas aeruginosa gene pfm affects the expression levels of type Ⅲ secretion system effectors[J]. Microbiology China, 2013, 40(8): 1462-1467. (in Chinese) 牟锐, 杜星, 王雪涵, 等. 铜绿假单胞菌pfm基因对三型分泌系统效应蛋白的影响[J]. 微生物学通报, 2013, 40(8): 1462-1467. |
| [106] |
Neidig A, Yeung ATY, Rosay T, et al. TypA is involved in virulence, antimicrobial resistance and biofilm formation in Pseudomonas aeruginosa[J]. BMC Microbiology, 2013, 13: 77. DOI:10.1186/1471-2180-13-77 |
| [107] |
McCarthy RR, Mooij MJ, Reen FJ, et al. A new regulator of pathogenicity (bvlR) is required for full virulence and tight microcolony formation in Pseudomonas aeruginosa[J]. Microbiology, 2014, 160(Pt 7): 1488-1500. |
| [108] |
Intile PJ, Balzer GJ, Wolfgang MC, et al. The RNA helicase DeaD stimulates ExsA translation to promote expression of the Pseudomonas aeruginosa type Ⅲ secretion system[J]. Journal of Bacteriology, 2015, 197(16): 2664-2674. DOI:10.1128/JB.00231-15 |
| [109] |
Wang BB, Li B, Liang Y, et al. Pleiotropic effects of temperature-regulated 2-OH-lauroytransferase (PA0011) on Pseudomonas aeruginosa antibiotic resistance, virulence and type Ⅲ secretion system[J]. Microbial Pathogenesis, 2016, 91: 5-17. DOI:10.1016/j.micpath.2015.11.003 |
| [110] |
Chen RH, Weng YD, Zhu F, et al. Polynucleotide phosphorylase regulates multiple virulence factors and the stabilities of small RNAs RsmY/Z in Pseudomonas aeruginosa[J]. Frontiers in Microbiology, 2016, 7: 247. |
| [111] |
Li M, Long YQ, Liu Y, et al. HigB of Pseudomonas aeruginosa enhances killing of phagocytes by up-regulating the type Ⅲ secretion system in ciprofloxacin induced persister cells[J]. Frontiers in Cellular and Infection Microbiology, 2016, 6: 125. |
| [112] |
Pérez-Gallego M, Torrens G, Castillo-Vera J, et al. Impact of AmpC derepression on fitness and virulence: the mechanism or the pathway?[J]. mBio, 2016, 7(5): e01783-16. |
| [113] |
Deng X, Li M, Pan XL, et al. Fis regulates type Ⅲ secretion system by influencing the transcription of exsA in Pseudomonas aeruginosa strain PA14[J]. Frontiers in Microbiology, 2017, 8: 669. |
| [114] |
Zhang YF, Han K, Chandler CE, et al. Probing the sRNA regulatory landscape of P. aeruginosa: post-transcriptional control of determinants of pathogenicity and antibiotic susceptibility[J]. Molecular Microbiology, 2017, 106(6): 919-937. |
| [115] |
Anantharajah A, Mingeot-Leclercq MP, Bambeke F. Targeting the type three secretion system in Pseudomonas aeruginosa[J]. Trends in Pharmacological Sciences, 2016, 37(9): 734-749. DOI:10.1016/j.tips.2016.05.011 |
| [116] |
Yamazaki A, Li J, Zeng Q, et al. Derivatives of plant phenolic compound affect the type Ⅲ secretion system of Pseudomonas aeruginosa via a GacS-GacA two-component signal transduction system[J]. Antimicrobial Agents and Chemotherapy, 2012, 56(1): 36-43. DOI:10.1128/AAC.00732-11 |
| [117] |
Shan JM, Wu XG, Yuan XC, et al. The influence of cinnamic acid derivatives on the type Ⅲ secretion system of Pseudomonas aeruginosa PAO1[J]. Microbiology China, 2014, 41(10): 2061-2069. (in Chinese) 单金明, 吴小刚, 袁小琛, 等. 肉桂酸衍生物对铜绿假单胞菌PAO1 Ⅲ型分泌系统的影响[J]. 微生物学通报, 2014, 41(10): 2061-2069. |
| [118] |
Grier MC, Garrity-Ryan LK, Bartlett VJ, et al. N-hydroxybenzimidazole inhibitors of ExsA MAR transcription factor in Pseudomonas aeruginosa: in vitro anti-virulence activity and metabolic stability[J]. Bioorganic & Medicinal Chemistry Letters, 2010, 20(11): 3380-3383. |
| [119] |
Marsden AE, King JM, Spies MA, et al. Inhibition of Pseudomonas aeruginosa ExsA DNA-binding activity by N-hydroxybenzimidazoles[J]. Antimicrobial Agents and Chemotherapy, 2016, 60(2): 766-776. DOI:10.1128/AAC.02242-15 |
| [120] |
Wang D, Zetterstrom CE, Gabrielsen M, et al. Identification of bacterial target proteins for the salicylidene acylhydrazide class of virulence-blocking compounds[J]. Journal of Biological Chemistry, 2011, 286(34): 29922-29931. DOI:10.1074/jbc.M111.233858 |
| [121] |
Enquist PA, Gylfe Å, Hägglund U, et al. Derivatives of 8-hydroxyquinoline — antibacterial agents that target intra- and extracellular Gram-negative pathogens[J]. Bioorganic & Medicinal Chemistry Letters, 2012, 22(10): 3550-3553. |
| [122] |
Felise HB, Nguyen HV, Pfuetzner RA, et al. An inhibitor of gram-negative bacterial virulence protein secretion[J]. Cell Host & Microbe, 2008, 4(4): 325-336. |
| [123] |
Aiello D, Williams JD, Majgier-Baranowska H, et al. Discovery and characterization of inhibitors of Pseudomonas aeruginosa type Ⅲ secretion[J]. Antimicrobial Agents and Chemotherapy, 2010, 54(5): 1988-1999. DOI:10.1128/AAC.01598-09 |
| [124] |
Zhang CF, Wu XG, Li Y, et al. Synthesis and bioactivity of novel inhibitors for type Ⅲ secretion system of Pseudomonas aeruginosa PAO1[J]. Chinese Journal of Organic Chemistry, 2013, 33(6): 1309-1318. (in Chinese) 张成芳, 吴小刚, 李燕, 等. 铜绿假单胞菌PAO1 Ⅲ型分泌系统新型抑制剂的合成和生物活性研究[J]. 有机化学, 2013, 33(6): 1309-1318. |
| [125] |
Gu LL, Wu XG, Yuan XC, et al. Regulation of type Ⅲ secretion system inhibitors on virulence factors of Pseudomonas aeruginosa PAO1[J]. Microbiology China, 2014, 41(11): 2318-2324. (in Chinese) 顾玲玲, 吴小刚, 袁小琛, 等. Ⅲ型分泌系统抑制剂对铜绿假单胞菌PAO1毒性因子的影响[J]. 微生物学通报, 2014, 41(11): 2318-2324. |
| [126] |
Frank DW, Vallis A, Wiener-Kronish JP, et al. Generation and characterization of a protective monoclonal antibody to Pseudomonas aeruginosa PcrV[J]. The Journal of Infectious Diseases, 2002, 186(1): 64-73. |
| [127] |
Imamura Y, Yanagihara K, Fukuda Y, et al. Effect of anti-PcrV antibody in a murine chronic airway Pseudomonas aeruginosa infection model[J]. European Respiratory Journal, 2007, 29(5): 965-968. |
| [128] |
Sawa T, Yahr TL, Ohara M, et al. Active and passive immunization with the Pseudomonas V antigen protects against type Ⅲ intoxication and lung injury[J]. Nature Medicine, 1999, 5(4): 392-398. DOI:10.1038/7391 |
| [129] |
Lynch SV, Flanagan JL, Sawa T, et al. Polymorphisms in the Pseudomonas aeruginosa type Ⅲ secretion protein, PcrV - implications for anti-PcrV immunotherapy[J]. Microbial Pathogenesis, 2010, 48(6): 197-204. DOI:10.1016/j.micpath.2010.02.008 |
| [130] |
DiGiandomenico A, Keller AE, Gao CH, et al. A multifunctional bispecific antibody protects against Pseudomonas aeruginosa[J]. Science Translational Medicine, 2014, 6(262): 262ra155. DOI:10.1126/scitranslmed.3009655 |
| [131] |
François B, Luyt CE, Dugard A, et al. Safety and pharmacokinetics of an anti-PcrV PEGylated monoclonal antibody fragment in mechanically ventilated patients colonized with Pseudomonas aeruginosa: a randomized, double-blind, placebo-controlled trial[J]. Critical Care Medicine, 2012, 40(8): 2320-2326. DOI:10.1097/CCM.0b013e31825334f6 |
| [132] |
Arnoldo A, Curak J, Kittanakom S, et al. Identification of small molecule inhibitors of Pseudomonas aeruginosa exoenzyme S using a yeast phenotypic screen[J]. PLoS Genetics, 2008, 4(2): e1000005. DOI:10.1371/journal.pgen.1000005 |
| [133] |
Lee VT, Pukatzki S, Sato H, et al. Pseudolipasin A is a specific inhibitor for phospholipase A2 activity of Pseudomonas aeruginosa cytotoxin ExoU[J]. Infection and Immunity, 2007, 75(3): 1089-1098. DOI:10.1128/IAI.01184-06 |
| [134] |
Kim D, Baek J, Song J, et al. Identification of arylsulfonamides as ExoU inhibitors[J]. Bioorganic & Medicinal Chemistry Letters, 2014, 24(16): 3823-3825. |