 2017, Vol. 44
2017, Vol. 44扩展功能
文章信息
- 曲文颖, 陈雷, 王光玉, 闫培生
- QU Wen-Ying, CHEN Lei, WANG Guang-Yu, YAN Pei-Sheng
- 海鞘与蓝藻的共生关系
- Symbiosis between ascidian and cyanobacteria
- 微生物学通报, 2017, 44(2): 458-464
- Microbiology China, 2017, 44(2): 458-464
- DOI: 10.13344/j.microbiol.china.160144
-
文章历史
- 收稿日期: 2016-02-18
- 接受日期: 2016-05-31
- 优先数字出版日期(www.cnki.net): 2016-06-06

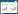




海鞘(Ascidian)属于脊索动物门尾索动物亚门海鞘纲,全世界约有3 000种[1]。海鞘结构简单,但有细胞分化。海鞘是滤食性的被囊动物,以水中细小的细菌、浮游生物和有机物颗粒为食,分解其中的有机物并排走代谢废物。
海鞘能在竞争异常激烈的海洋生境中繁衍生息,主要依赖于其强大的化学防御机制,能够产生大量结构新颖的天然产物。在过去约40年的时间里,已从海鞘中分离到1 080种天然产物,许多天然产物可作为药物先导化合物,具有抗肿瘤、抗微生物、抗病毒和免疫调节等作用,是目前已经发现天然产物最多的海洋动物之一[2-3]。有直接和间接证据表明,海鞘中的大部分活性物质是由共生细菌产生的[4-5]。如Et-743 (商品名Yondelis)是四氢异喹啉生物碱类化合物,最初是从加勒比海鞘(Ecteinascidia turbinata)中分离得到的,后来被证明是由其共生细菌(Candidatus Endoecteinascidia frumentensis)产生[6]。2007年Et-743通过了欧盟认证,用于治疗软组织肉瘤,2009年用于治疗卵巢癌[5]。Patellazole是从海鞘Lissoclinum patella中分离到的聚酮类化合物,具有良好的细胞毒活性,其真正来源是海鞘的共生细菌Candidatus Endolissoclinum faulkneri[7]。
共生(Symbiosis)是1879年德国生物学家Anton de Bary提出的,是指两生物体之间生活在一起的相互作用。动物、植物、菌类以及三者中任意 两者之间都可能形成共生关系。在海鞘与蓝藻的共生关系中主要是互利共生(Mutualism),即一方为另一方提供有利于生存的帮助,同时也获得对方的帮助。依照共生关系中两者个体大小的区别,又可分为宿主和共生体。宿主(Host)通常指共生关系中个体较大的成员,较小者称为共生体(Symbiont)。
本文以海鞘与蓝藻共生关系的典型实例为代表,介绍了两者在共生关系中可能存在的生理功能,为进一步开发利用海洋生物之间的共生关系提供研究基础。
1 蓝藻在海鞘中的存在1931年,Hastings认为海鞘中存在虫黄藻。1975年,Newcomb和Pugh鉴定了海鞘中的藻类不是虫黄藻而是蓝藻。同年,Lewin发现了几种只与Didemnid海鞘共生的独特藻类,其含有叶绿素a和b,但缺乏藻胆色素,有的种类会有堆积在一起的类囊体。1977年,Whatley发现与Didemnid海鞘共生的藻类没有核膜或膜性细胞器,细胞壁为 四层,且肽聚糖结构复杂。同年,Lewin将与Didemnid海鞘共生的藻作为藻类分类中的一个新属,原绿藻属(Prochloron)。1982年,Kott发现有20种海鞘都存在与藻类共生的现象,其中17种海鞘都与原绿藻属的蓝藻共生,其余3种海鞘(Trididemnum solidum、T. cyanophorum和Didemnum viride)与Synechocystis属的蓝藻共生[8]。
在海鞘的表面、泄殖腔及被膜中,共生的蓝藻形成一层易于辨识的生物膜。用荧光显微镜、荧光原位杂交(Fluorescence in situ hybridization,FISH)和酶联荧光原位杂交(Catalyzed reporter deposition fluorescent in situ hybridization,CARD-FISH)等技术可以定位蓝藻在海鞘中的具体位置。原绿藻在海鞘中的存在位置可能与维持共生关系有关。有人认为原绿藻存在于泄殖腔中竞争力强,可以阻碍其他蓝藻的进入,最有利于共生。为保证共生关系的稳定,海鞘中的原绿藻随着海鞘的繁殖进行世代传递,由于原绿藻本身没有运动能力,它们只能进行被动的水平或垂直传递。对于原绿藻在海鞘间的水平传递来说,不同的原绿藻在海鞘中有不同的分布方式,它们可以通过改变自身的形态及分布来适应新的宿主;对于垂直传递来说,原绿藻就没有这种适应性的改变[9]。
原绿藻在海鞘间进行世代传递时有两种方式。第一种:要求其必需在海鞘胚胎的后期存在于海鞘的泄殖腔中,然后海鞘的胚胎被转移到海鞘被膜上,胚胎形成蝌蚪型的幼体进入泄殖腔,原绿藻细胞黏附在幼体躯干的后半段(因前半段有膜包裹,无黏性,不能黏附原绿藻),最终与幼体一起离开泄殖腔,发育成为新的海鞘。第二种:海鞘被膜中未成熟的幼体向后伸出穗状结构,原绿藻黏附到该穗状结构上进入泄殖腔中,幼体离开母体时,幼体表面的被膜在幼体后半部分形成一个袋状结构,到海鞘变形期时,袋状结构形成泄殖腔。原绿藻的两种传递方式都发生在海鞘胚胎时期。有些海鞘中含有多种蓝藻,这些蓝藻对于海鞘来说不是必需的,在世代传递的过程中,这些蓝藻可能在被膜中发生竞争,产生排斥、分离或共生的现象[9]。
2 蓝藻共生体的生理功能与海鞘共生的细菌中,最有代表性的主要是细菌界(Bacteria)蓝藻门(Cyanobacteria)的原绿藻属(Prochloron)。目前原绿藻属(Prochloron)只有一个种Prochloron didemni,因此本文提到的各研究中使用的P. didemni都只用原绿藻的名字代替。在藻类生物中,蓝藻是最简单、最原始的单细胞生物,有些蓝藻可以进行碳固定,有些蓝藻还可以进行氮固定。研究最为广泛的是热带水域的Didemnid海鞘与原绿藻的共生。Didemnid海鞘通常是生态系统的主要组分,潜在影响着珊瑚礁的结构[10]。到目前为止还未发现有自由生活的原绿藻,这说明原绿藻必需依赖海鞘才能生存[11]。原绿藻在碳固定、氮的循环利用以及代谢产物生成方面对宿主起到一定的作用。
2.1 蓝藻在海鞘中进行碳固定1942年,Tokioka首次发现海鞘(Diplosoma virens)中的藻类共生体可以通过光合作用产生氧气,藻类的新陈代谢对宿主海鞘产生有利影响。1978年,Akazawa等推断海鞘D. virens中的藻类可能通过C3途径固定CO2[12]。1980年,科学家在水族箱中培养含有藻类的玻璃海鞘(Ciona intestinalis),发现海鞘在全透光下比在半透光下生长得好,并且在黑暗条件中一定会死亡,这为探讨两者的共生关系提供了间接的证据[13]。
对蓝藻在海鞘中进行碳固定的研究最有效的方法是同位素示踪法和基因组测序法。同位素示踪法是用同位素14C标记水中的碳,不仅可以检测到原绿藻光合产物的量,还可以观察到光合产物在两者之间的转运途径,从而说明其共生关系。通过对蓝藻基因序列及其功能的研究,不但可以发现蓝藻中各种蛋白质结构因共生关系而发生的变化,而且为阐述固碳机制和共生关系提供新的证据。
在研究海鞘与蓝藻的共生关系中常使用的 两种海鞘宿主是Diplosoma sp.和L. patella,因为从中很容易分离到原绿藻。用14C标记NaH14CO3的海水培养这两种海鞘或用这种海水配制的培养基培养原绿藻,结果都表明海鞘体内的藻类在光下可以通过光合作用利用海水中的HCO3-合成有机物。海鞘可以利用蓝藻合成的光合产物合成自身所需要的重要物质,如脂质、核酸和蛋白质。向含有原绿藻的海鞘(Trididemnum cyclops)供给用14C标记的醋酸,并且用太阳光(800 lx)照射150 min,发现醋酸在海鞘体内转化的产物中有70%为极性脂质(推测是磷脂),10%为脂肪酸,6%为甾醇,13%为蜡和烃,以及不足1%的甘油三酯。这些放射性碳还可能参与磷酸烯醇式丙酮酸羧化酶的代谢,此酶涉及到糖的代谢过程[12]。原绿藻的基因组测序也表明,原绿藻可以合成海鞘宿主体内大多数的脂类物质,包括一些潜在的生物燃料分子[10]。另外,不同种类的海鞘对蓝藻提供有机碳的依赖程度不同,如与海鞘(Lissoclinum voeltzkowi)共生的原绿藻几乎可以为海鞘提供所需的全部碳源,与海鞘(Didemnum molle)共生的原绿藻只能为其提供60%的有机碳,余下的需要通过海鞘摄食获得[14]。这些都说明原绿藻的固碳作用对于海鞘很重要,有些海鞘就是依靠这些共生体才能在热带海域贫营养的条件下得以生存(图 1)。

|
| 图 1 海鞘与原绿藻的共生关系 Figure 1 Symbiotic relationship between ascidian and Prochloron |
|
|
为了进一步证明蓝藻的碳固定功能,现在已经在基因组水平上对其进行了研究。对与海鞘L. patella共生的原绿藻的光合作用相关基因序列筛查表明,原绿藻含有一套完整的光合作用基因。但原绿藻为了适应其生活的贫营养环境,基本结构产生一些重要的变化,如它虽无叶绿体,但在电镜下可见细胞质中有很多光合片层,即类囊体,各种光合色素均附于其上,含有膜内在蛋白的扩展家族,以及含有叶绿素a和b的光采集复合物蛋白等。叶绿素a和b扩大了蓝藻对可见光区域中蓝光和红光的捕获能力[10],这些都是原绿藻进行光合作用的基础。再者,由于原绿藻与海鞘共生,所以动物体可以持续供给原绿藻CO2,因此原绿藻中负责CO2运输的基因相比其他种类的蓝藻要小很多。
2.2 蓝藻在海鞘中进行氮循环利用在贫营养的热带水域,氮的来源是令人费解的。这里一年中的大部分时间总氮环境一般小于0.2 μmol/L。在海鞘和蓝藻共生的栖息地,其海水非常清澈,颗粒物很少,海水中颗粒物的有机氮浓度只有0.5-5.0 μmol/L[15]。通过对2种海鞘L. voeltzkowi和D. molle碳氮营养的研究发现,原绿藻对氮素的需求量远远高于周围环境能够提供的量;海鞘通过周围环境获取的氮远远低于海鞘向外界排出的氮,即摄取的氮不足以满足共生系统生存的需要。因此,海鞘与共生的蓝藻如何有效经济地利用氮源以应对生存威胁就至关重要了[14]。
2011年Donia通过对二段海鞘(L. patelle)基因组与原绿藻基因组的测定发现,在原绿藻的基因组中没有发现氮固定所必需的固氮酶基因nifD和nifH,因此原绿藻不能直接固定大气中的氮。继续对该种海鞘中存在的其他微生物的宏基因组进行测定,发现海鞘中其他微生物也没有固氮酶的基因,这表明海鞘体内不能进行固氮作用[10]。
原绿藻可利用海水中的无机氮,另外海鞘摄取的食物中的氮经体内代谢后产生的含氮代谢废物也可供原绿藻使用,原绿藻可以将这些含氮化合物进行转化再次循环使用(图 1)。原绿藻的基因序列中含有可以将氨转换成谷氨酰胺的基因,可以将光呼吸产生的NH4+和硝酸盐还原产生的NH4+通过谷氨酰胺合成酶/谷氨酸合成酶转变成谷氨酰胺。原绿藻可以进行有效的氮素循环利用是海鞘与原绿藻共生体系中最重要的氮的来源[10]。
2.3 蓝藻在海鞘中产生次级代谢产物海鞘大多身体柔软,极易被捕食,需要化学防御来保护其生存。现在已经从海鞘中分离出 1 000多种结构不同的天然产物,其中有很多可能是由其共生微生物产生的[10]。
从一些蓝藻中克隆出非核糖体肽合成酶(Nonribosomal peptides synthetase,NRPS)基因,证明蓝藻能够产生一些次级代谢产物[16]。它们可积累在细胞内,也可排到外环境中。代谢产物大多具有生物活性,在蓝藻与海鞘的共生过程中,有的代谢产物可帮助海鞘生存。在Didemnid海鞘的提取物中发现了Patellamide类的环肽,接着从与之共生的原绿藻中找到了此环肽的生物合成基因,并在大肠杆菌(Escherichia coli)中进行了整个合成途径基因的异源表达以确定环肽的生理功能。这种环肽可以螯合铜、锌等金属离子,对海鞘具有解毒作用;但环肽也有细胞毒性,可以抑制海鞘摄食[17]。
海鞘中的共生蓝藻可以产生多种多样的次级代谢产物。对L. patella海鞘中的原绿藻基因组进行研究,虽然4个L. patella海鞘的采集地点相距较远而且其体内化合物的成分差异较大,但是4个海鞘体内的原绿藻基因组序列中负责初级代谢的功能基因几乎是完全相同的,只有一部分负责次级代谢的基因不同。原绿藻产生不同次级代谢产物的机制可能有3种:(1) 存在不同的生物合成基因簇,可以合成不同的次级代谢产物;(2) 生物合成途径间进行同源重组,同源基因相互协作一起完成次级代谢产物的合成。同源基因可能编码不同的代谢产物,一种基因也可能编码多种代谢产物;(3) 前体肽的高突变率。由于合成一些次级代谢产物的生物合成酶具有宽泛的底物特异性,所以可以产生多种次级代谢产物[2]。
共生体产生的化合物能够使宿主在不良环境中进行化学防御,那么海鞘是如何将能产生特定化合物的微生物选作自己的共生体,其机理尚待研究[18]。另外,一种海鞘体内可能有多株与之共生的原绿藻,每株原绿藻合成Patellamide环肽类细胞毒素的途径不同,因此能够产生多种细胞毒素,可以用于海鞘的化学防御(图 1)[9]。
3 海鞘对蓝藻共生体的作用 3.1 海鞘对共生体的UV保护作用海鞘多生活在海洋的浅水区域。太阳光中的紫外线可以穿透超过16 m深的海水,且在浅水区域的紫外线强度高于其他海洋环境,对生物体具有严重的杀伤作用。海洋动物暴露在紫外线辐射下可能造成DNA损伤,导致基因突变率增加[19]。因此需要探讨海鞘与共生体在很强的紫外线辐射下的生存机制。
20世纪90年代,科学家发现紫外线可以通过氧中毒的形式间接杀伤生物。然而有些生物呈现出对高强度光的一种适应性,来减少对光合作用的抑制。这种对紫外线的保护机制可能是通过合成紫外吸收化合物来过滤掉一些有害波长的光,或者利用某些酶来抵御氧的毒害作用[20]。
海鞘对共生体具有防紫外线辐射的保护作用(图 1)。虽然海鞘表面的被膜能透过光,但这种厚厚的胶质或革质的被膜可以起到一定的保护作用,再加上海鞘能够合成几种紫外吸收化合物——类菌孢素氨基酸(Mycosporine-like amino acids,MAAs)[20-21],如shinorine、菌胞素-甘氨酸(mycosporine-glycine)和palythine,可以实现对原绿藻的抗紫外线的防护。虽然原绿藻自身也含有编码MAAs的合成基因,具有合成MAAs的能力,但是其合成的数量较少不能满足需求,所以还需要宿主海鞘的保护作用[20]。
3.2 海鞘为共生体的氮循环利用提供保障在海鞘与原绿藻的共生系统中,海鞘可以通过摄食获取环境中的有机氮,经体内代谢后将产生的含氮代谢废物提供给共生体原绿藻,通过原绿藻的转化作用使氮素在共生系统中有效的循环使用。因此海鞘为整个共生系统的氮素营养提供一定的来源和保障(图 1)。
4 展望海鞘与原绿藻的共生关系是海洋动物与藻类共生关系的典型代表,虽然在近20年的研究中,已经对两者在共生关系中可能存在的生理功能进行了研究,但是还有很多机理没有被解释清楚,或者很多还仅仅是猜测而缺乏证据支持。因此需要通过化学、微生物学和分子生物学等学科间的合作进一步加深对共生关系的研究,为进一步开发利用海洋生物之间的共生关系提供研究基础。
4.1 蓝藻共生体的体外培养原绿藻必需依赖于海鞘才能生存,目前还未发现有可独立存活的原绿藻。这对于研究海鞘与原绿藻之间的共生关系、生理功能、代谢产物的合成途径等存在一定的困难。因此如何实现原绿藻的体外培养将是一项非常有意义的研究。
体外培养原绿藻的关键可能首先要深入了解原绿藻成为海鞘共生体的原因。然后可借助现代提纯和分析技术,研究海鞘的摄食和体内环境,对海鞘分泌物和排泄物进行全面系统的分析,找出分泌物和排泄物与共生蓝藻生长发育间的关系;可借助气质联用仪或液质联用仪等仪器对物质组分进行分析。在此基础上进行原绿藻培养基的配制,并改变一些传统的前处理和平板培养的方法,比如研磨可能会损伤藻细胞或者纱布过滤时可能会漏掉某些藻类等,改进培养方法可能会实现某些原绿藻的人工培养。
关于与海鞘共生的原绿藻的分离,可以借鉴海绵共生蓝藻的分离方法。海绵(Dysidea herbacea)的共生蓝藻(Oscillatoria spongeliae)占据了海绵组织的20%,可以使用流式细胞仪从海绵细胞中分离出蓝藻[22]。方便快捷地从海鞘中获得很多原绿藻细胞对于今后原绿藻的体外培养等研究将起着重要的作用。
4.2 蓝藻代谢产物生理功能的研究与海鞘共生的蓝藻可以产生很多代谢产物,这些代谢产物在体外具有抗肿瘤、抗疟、抗菌等生物活性。现在猜测这些代谢产物对于海鞘的化学防御有一定的作用,今后需要提供一定的证据来加以证明。如可通过在已添加病毒等不利于海鞘生长的环境中培养海鞘,一段时间后检测海鞘体内的某些物质含量是否增加,含量增加的物质可能就是具有抗病毒功能的代谢产物,而后利用基因组学方法分别检测海鞘和蓝藻共生体,找到合成该代谢产物的基因,明确代谢产物的真正来源,进而研究其在共生关系中的作用。某些可被用作药物的代谢物由不可培养的共生体产生,可将产生代谢物的生物合成基因转入可培养的细菌中,从而就可以持续获得该代谢物[22]。另外,以后还应该研究宿主-微生物共生关系的生态学意义,例如天然产物在建立和维护共生关系平衡过程中起到的作用[23]。
4.3 共生关系的起源和进化的研究现在虽然已经能够通过一些技术确定蓝藻在海鞘中的具体位置,以及对蓝藻在海鞘世代间的垂直传递给出一定的解释。但是海鞘和蓝藻之间共生的起源是如何发生的?海鞘如何选择不同的蓝藻?海鞘为了能在世代间传递蓝藻,其自身的结构和基因发生了怎样的改变?蓝藻在海鞘间如何进行水平传递?所有这些问题都需要通过遗传学和生态学上的研究,找出相互作用的共生体如何进化的证据。
在研究海鞘与蓝藻共生关系的起源和进化时往往会遇到非专一性共生的问题,即与原绿藻共生的海鞘种类不具有专一性。虽然与原绿藻共生的海鞘种类有很大一部分属于Didemnidae科(也称为Didemnid海鞘),但是Didemnidae科的海鞘中也有一些不与藻类共生的海鞘种类。另外,每个Didemnid海鞘的属中,既包括与原绿藻共生的海鞘种,又包括不与原绿藻共生的海鞘种[10]。虽然一直以来Didemnidae科是单系(Monophyly)的,即来源于同一个祖先的说法被广泛接受,但是用分子生物学的方法对这个科的各个属间进行系统发育关系的研究结果却无法支持这个结论[24]。今后还需要对海鞘和共生体两者相结合进行更为深入的研究,才能更好地理解它们之间的系统进化关系。
综上所述,对海鞘与蓝藻共生关系的研究将有助于理解生物进化过程,以及为进一步开发利用海洋生物之间的共生关系提供研究基础。
| [1] | da Silva Oliveira FA, Colares GB, Hissa DC, et al. Microbial epibionts of the colonial ascidians Didemnum galacteum and Cystodytes sp[J]. Symbiosis 2013, 59(1) : 57–63. DOI:10.1007/s13199-012-0210-2 |
| [2] | Schmidt EW, Donia MS, McIntosh JA, et al. Origin and variation of tunicate secondary metabolites[J]. Journal of Natural Products 2012, 75(2) : 295–304. DOI:10.1021/np200665k |
| [3] | Blunt JW, Copp BR, Munro MHG, et al. Marine natural products[J]. Natural Product Reports 2011, 28(2) : 196–268. DOI:10.1039/C005001F |
| [4] | Chen L, Fu CM, Wang GY. Microbial diversity associated with ascidians: a review of research methods and application[J]. Symbiosis 2017, 71(1) : 19–26. DOI:10.1007/s13199-016-0398-7 |
| [5] | Fu CM. Bacterial diversity associated with two species of ascidians and screening of cytotoxic activity of the strains[D]. Weihai: Master’s Thesis of Harbin Institute of Technology, 2015 (in Chinese) 符常明. 两种海鞘相关可培养细菌多样性和抗肿瘤活性筛选[D]. 威海: 哈尔滨工业大学硕士学位论文, 2015 |
| [6] | Rath CM, Janto B, Earl J. Meta-omic characterization of the marine invertebrate microbial consortium that produces the chemotherapeutic natural product ET-743[J]. ACS Chemical Biology 2011, 6(11) : 1244–1256. DOI:10.1021/cb200244t |
| [7] | Kwan JC, Schmidt EW. Bacterial endosymbiosis in a chordate host: long-term co-evolution and conservation of secondary metabolism[J]. PLoS One 2013, 8(12) : e80822. DOI:10.1371/journal.pone.0080822 |
| [8] | Olson RR. Photoadaptations of the Caribbean colonial ascidian-cyanophyte symbiosis Trididemnum solidum[J]. Biological Bulletin 1986, 170(1) : 62–74. DOI:10.2307/1541381 |
| [9] | Hirose E. Ascidian photosymbiosis: diversity of cyanobacterial transmission during embryogenesis[J]. Genesis 2015, 53(1) : 121–131. DOI:10.1002/dvg.v53.1 |
| [10] | Donia MS, Fricke WF, Partensky F, et al. Complex microbiome underlying secondary and primary metabolism in the tunicate-Prochloron symbiosis[J]. Proceedings of the National Academy of Sciences of the United States of America 2011, 108(51) : E1423–E1432. DOI:10.1073/pnas.1111712108 |
| [11] | Hirose E, Maruyama T. What are the benefits in the ascidian-Prochloron symbiosis[J]. Endocytobiosis and Cell Research 2004, 15(1) : 51–62. |
| [12] | Pardy RL, Lewin RA. Colonial ascidians with prochlorophyte symbionts: evidence for translocation of metabolites from alga to host[J]. Bulletin of Marine Science 1981, 31(4) : 817–823. |
| [13] | de Leo G, Patricolo E. Blue-green algalike cells associated with the tunic of Ciona intestinalis L[J]. Cell and Tissue Research 1980, 212(1) : 91–98. DOI:10.1007/BF00234036 |
| [14] | Koike I, Yamamuro M, Pollard PC. Carbon and nitrogen budgets of two ascidians and their symbiont, Prochloron, in a tropical seagrass meadow[J]. Australian Journal of Marine and Freshwater Research 1993, 44(1) : 173–182. |
| [15] | Paerl HW. N2 fixation (nitrogenase activity) attributable to a specific Prochloron (Prochlorophyta)-ascidian association in Palau, Micronesia[J]. Marine Biology 1984, 81(3) : 251–254. DOI:10.1007/BF00393219 |
| [16] | Kleigrewe K, Gerwick L, Sherman DH, et al. Unique marine derived cyanobacterial biosynthetic genes for chemical diversity[J]. Natural Product Reports 2016, 33(2) : 348–364. DOI:10.1039/C5NP00097A |
| [17] | Schmidt EW, Nelson JT, Rasko DA. Patellamide A and C biosynthesis by a microcin-like pathway in Prochloron didemni, the cyanobacterial symbiont of Lissoclinum patella[J]. Proceedings of the National Academy of Sciences of the United States of America 2005, 102(20) : 7315–7320. DOI:10.1073/pnas.0501424102 |
| [18] | Schmidt EW. The secret to a successful relationship: lasting chemistry between ascidians and their symbiotic bacteria[J]. Invertebrate Biology 2015, 134(1) : 88–102. DOI:10.1111/ivb.2015.134.issue-1 |
| [19] | Svanfeldt K, Lundqvist L, Rabinowitz C, et al. Repair of UV-induced DNA damage in shallow water colonial marine species[J]. Journal of Experimental Marine Biology and Ecology 2014, 452 : 40–46. DOI:10.1016/j.jembe.2013.12.003 |
| [20] | Dionisio-Sese ML, Ishikura M, Maruyama T, et al. UV-absorbing substances in the tunic of a colonial ascidian protect its symbiont, Prochloron sp., from damage by UV-B radiation[J]. Marine Biology 1997, 128(3) : 455–461. DOI:10.1007/s002270050112 |
| [21] | Lesser MP, Stochaj WR. Photoadaptation and protection against active forms of oxygen in the symbiotic procaryote Prochloron sp. and its ascidian host[J]. Applied and Environmental Microbiology 1990, 56(6) : 153–1535. |
| [22] | Piel J. Metabolites from symbiotic bacteria[J]. Natural Product Reports 2004, 21(4) : 519–538. DOI:10.1039/b310175b |
| [23] | Konig GM, Kehraus S, Seibert SF, et al. Natural products from marine organisms and their associated microbes[J]. Chembiochem 2006, 7(2) : 229–238. DOI:10.1002/cbic.v7:2 |
| [24] | Hirose M, Hirose E. Photosymbiotic ascidians from oceanic islands in the tropical Pacific as candidates of long-dispersal species: morphological and genetic identification of the species[J]. Aquatic Invasions 2013, 8(3) : 271–280. DOI:10.3391/ai |